Blaming Our Genes: The Heritability of Behavior

It’s easy to accept that human disorders such as phenylketonuria or cystic fibrosis or Huntington’s disease have a wholly genetic basis. And you likely have no problem believing that your risk of being afflicted with an illness such as heart disease, diabetes, or colon cancer is influenced by your personal DNA code. The question of heredity becomes more complicated, however, when we consider complex behaviors.
Is your chance of having a sunny disposition affected by your genes? How about if you are a pessimist — is pessimism an inherited trait? What if you’re an early-morning or a late-night person, or compulsively neat, or emotionally unable to connect with others — could genetic differences contribute to those traits? How much of our temperament is due to our genes? How much of our intelligence — our ability to learn and remember, to acquire language, to read and spell — is a function of our DNA code? And what about psychiatric disorders such as schizophrenia, bipolar disorder, and depression? Are those due to our circumstances, or to our genetic constitution?
The answer is: both. Nature and nurture, our genetic endowment and our life situation and experiences, determine our behavior. But we suspect that the contribution from genetics may be more than most of us imagine.
Consider, for example, Susan Middlebrook, of Colchester, Vermont. She usually gets seven or eight hours of sleep each night, but rather than going to bed at 11:30 p.m., after the late-night news, and getting up at around 7 a.m., Middlebrook goes to bed early, very early . . . like at 6:30 p.m. And between 1:30 and 3 a.m., just when you’re deep in your dreams, Middlebrook is raring to get up and go. “The net result is you can feel very isolated,” she told National Geographic News in 2005. “Who wants to party at three in the morning? Nobody I know, and I’m not headed to the local bar to see who’s there.” Instead, Middlebrook gets started on her morning chores long before your dreams are over.

Middlebrook suffers from familial advanced sleep phase syndrome (FASPS), the consequence of which is that her sleep patterns are way out of sync with the norm. To establish the norm, and the abnormal exceptions, sleep researchers ask people a series of questions about their sleep habits: At what time of day do you feel your best? How easy is it for you to get up in the morning? At what time would you go to bed if you were completely free to plan your day? At what time in the evening do you feel drowsy and begin to doze? Do you consider yourself a “morning” or an “evening” person? (The questionnaire was developed by a British scientist, James Horne, and a Swedish scientist, Olov Östberg.) Answers to these and other questions produce a score that lies on a scale running from extreme “eveningness” to extreme “morningness”; most of us fall somewhere in the middle of a broad bell-shaped curve of scores, but FASPS patients lie at the far “morningness” end of the scale, scoring higher than more than 99 percent of other respondents.
What makes Middlebrook’s syndrome so interesting is that it’s the result of an abnormality in her biological clock, known as the circadian rhythm; (“circadian” means, literally, the cycle of one complete day, from the Latin circa- , “cycle,” and dies , “day”). This circadian clock controls not just sleeping and waking but also metabolic, physiological and behavioral processes such as heart rate, hormone levels, blood pressure, mood, and alertness. Our biological clocks are reset each day by exposure to sunlight, thus keeping us in synchrony with our surroundings. But Middlebrook’s internal clock runs on a faster cycle, advanced by about four hours relative to most everyone else’s, so that she is ready for bed when her neighbors are ready to sit down to dinner.
Middlebrook’s sleep-wake cycle may seem unusual to you — it is far outside the statistical norm — but around the Middlebrook home it’s about as unusual as oatmeal for breakfast: two of her three sisters, one of her parents, and her own child all keep the same odd hours. This syndrome is clearly due to a variation present in the personal DNA code of these family members (the word “familial” in the name of the syndrome might have tipped you off to this), and not to any environmental cause.
Middlebrook’s sleep-wake cycle may seem unusual to you, but two of her three sisters, one of her parents, and her own child all keep the same odd hours.
Although FASPS affects only about three people in one thousand, its genetic basis was well worth deciphering. Findings about this syndrome may help to explain other more common sleeping disorders such as insomnia and narcolepsy, or lead to treatments for jet lag or seasonal affective disorder (SAD, in which the moods of sufferers are strongly affected by lack of sunlight in the winter), and they may provide health support and guidance for millions of workers who work the nightshift. Understanding human circadian rhythms may even enable drug treatments to be timed for better effect, or suggest ways to reduce nighttime auto accidents.
Now consider Jean, a 24-year old nurse who became obsessed with cleaning and washing. As related by Peter McGuffin and David Mawson, two psychiatrists in London to whom Jean was eventually referred, each day she would wash her hands 60 to 80 times and spend 12 hours disinfecting her house; she went through twenty liters of disinfectant a week. She made her husband stop his sports activity because it brought dirt into the house, and she gave up sexual activity because it could cause contamination. When Jean was admitted to the hospital, her hands were “roughened, red, cracked and bleeding” from the constant washing.
Jean’s identical twin, Jill, a social worker, lived apart from her sister and rarely saw her. Yet at the age of 22, Jill began displaying behavior similar to Jean’s. As soon as a meal was over she felt compelled to immediately wash the dishes and silverware, or else she would become severely anxious. Moreover, “the washing of the dishes and utensils had to proceed in a specific order, and failure to comply with the routine, or an attempt by others to relieve her of the task, provoked great discomfort.” Jill established other washing rituals, and despite her best efforts to stop carrying them out, she spent increasing amounts of time on them, to the detriment of her social life and her work.
Both twins had normal childhoods, had been outgoing children, and were academically accomplished. None of their family members had received treatment for a psychiatric disorder, although their father was “fastidiously neat and orderly in his habits.” The twins attended different universities; Jean obtained a degree in physics, Jill in history. Neither of them displayed any signs of mental illness before their obsessive-compulsive behaviors began. In fact, each of them learned of the other’s similar problem only after Jean began treatment with the drug clomipramine, which together with behavior therapy alleviated her symptoms enough to allow her to return to nursing. Jill was not treated, but experienced a spontaneous remission of her symptoms after more than a year.
What caused these identical twins to develop a similar disorder at a similar time in their lives? Was it something about the way their parents brought them up that led to their symptoms years later? Was it some food they ate, or some toxin they were exposed to as children? Or was it because their personal DNA code is identical, and some combination of variations in the genes they both inherited caused them to develop an obsessive-compulsive disorder in their early 20s? Or did the disorder develop because of a complex interplay of these factors?
Consider, finally, Lewis, the son of a well-to-do couple in Pittsburgh. Lewis looked perfect at birth and appeared to be developing normally. As detailed by his mother, at 18 months Lewis was almost saying words, but they weren’t the typical words that most toddlers begin with, such as “Mommy” and “Daddy.” Instead he babbled nonsensically. And he didn’t respond to the usual games that toddlers enjoy, such as peek-a-boo or ring-around-the-rosie. At two years of age, he had no interest in some toys but was obsessed with others, and when he played with these he was oblivious to everything going on around him. He loved to climb and to swing and to bounce on a trampoline, which he did without fear. Yet sometimes he was afraid to step off a rug onto a hardwood floor, reacting as if the next step would be off a cliff. At night, Lewis took off all his clothes and slept on the floor wrapped in a blanket and surrounded by toy soldiers and matchbox cars. Lewis also had tantrums, during which he lay on the floor screaming, unable to indicate his problem.
At three he had yet to acquire any language and had little interest in communicating with his family. He babbled only to his toy soldiers. His doctors were no longer able to dismiss his problems as simply those of a “difficult” child or one in the throes of the “terrible twos.” Lewis underwent a series of tests and the doctors concluded he had an autism spectrum disorder.
The diagnosis led to a variety of publicly and privately funded treatments, including speech and occupational therapy. Lewis made remarkable progress: He developed some language ability, and his behavior calmed down. He could hold his mother’s hand, play peek-a-boo with her and kiss her good night, and occasionally even make eye contact. When he read his favorite book, Eric Carle’s “The Very Hungry Caterpillar,” he said every word at the right time and mimicked the slurping sound of the caterpillar eating. But he still could not carry on a conversation. Lewis entered a special-education preschool class, with the hope and expectation that in a few years he could be integrated into a typical school class.
Autism is a complex developmental disorder that appears within the first three years of life. Its symptoms, which can range from mild to severe, are characterized by a lack of emotional contact with others, difficulty with verbal and nonverbal communication, and a restricted range of activities and interests. Children are born with autism or with the potential to develop it; they do not acquire the condition because of bad parenting. A fascinating feature of the chromosomes of some autistic children is that large chunks of DNA — millions or tens of millions of base-pairs — are found duplicated in some children and lost altogether in others. These regions can contain dozens of genes. The analysis of the personal DNA codes of these children may help pinpoint the specific genetic changes responsible for this complex and debilitating disease.
When a human population is measured for any characteristic, say height or weight, or blood pressure or cholesterol level, the measurements reveal a continuous spectrum between two extremes.
It’s no different for a complex behavior, such as how we deal with stress. Even very young babies display a broad range of responses, from those who cry easily to those who seem unbothered by almost any challenge. Similar patterns are observed in the extent or degree to which children are active or sedentary, whether they persist at pursuing a task or lose their focus, or the extent of their introversion or extroversion. Among adults, the amounts of alcohol, nicotine, or drugs that are consumed show the same broad distributions. Some part of these traits is likely due to genetics, and some is surely due to environmental components. But how much is due to each?
The measure of the amount of the genetic effect is termed “heritability.” A trait or behavior that is wholly due to genetics has 100 percent heritability; one that is wholly due to the environment has zero heritability.
The environmental component could be due to factors shared among family members, such as the home environment, the food preferences of the family, or the level of pollution in the town where the family resides. Or it could be due to factors that are not shared, such as the specific set of friends or teachers of each family member, or unusual life events such as accidents, or diseases unique to each individual.
How do we measure, or quantify, heritability? It’s actually pretty easy. All that is needed is a set of individuals whose genetic relatedness is known, and a measurement of some trait or condition for each individual.
The easiest of these studies to understand exploits the genetic similarity of twins. The personal DNA codes of identical twins are identical, because they develop from a single fertilized egg. That is, the DNA sequence of each gene in one twin is identical to its sequence in the other twin. Fraternal twins also shared the same womb at the same time, but have the same version of only about half of their genes (like any siblings), so the genetic contribution to a trait is twice as great in identical twins as it is in fraternal twins. Thus, the more similar a trait is in identical twins as compared to fraternal ones, the greater the genetic contribution to it is likely to be.
A large number of studies suggest a heritability for intelligence of around 50 percent; some studies put this number at greater than 80 percent.
Another type of study compares siblings from the same parents (who have the same version of about half of their genes), to adopted siblings who are not genetically related but share the same surroundings. A genetic influence on a trait is apparent when biological siblings are more similar than adopted ones; an environmental influence is obvious when adopted and thus genetically unrelated siblings resemble each other more than they do other unrelated people who grew up in other families.
A third type of analysis uses family genetic studies that compare how often a disease occurs in a family in which one member is known to be affected to how often it occurs in the general population. Diseases with a genetic basis occur more frequently in members of a family that has one case — in other words, genetic diseases run in families.
With these ways to quantify heritability, let’s examine some human traits. We’ll start with intelligence — a particularly thorny issue. We’d like to believe that a roomful of books, Mozart playing in the background, and a nurturing set of parents will put any infant on the road to a Nobel Prize. Those conditions can’t hurt, but they can only do so much: A large number of studies suggest a heritability for intelligence of around 50 percent; some studies put this number at greater than 80 percent.
A Swedish twin registry has information on 25,000 same-sex twins born over a period of about 70 years. Nancy Pedersen and her colleagues at the Karolinska Institute in Stockholm studied about 300 of these twin pairs, including identical twins reared apart, identical twins reared together, fraternal twins reared apart, and fraternal twins reared together. They found that intelligence scores of identical twins who grew up apart (most having been separated by the time they were two years old) correlated much more closely than the intelligence scores of fraternal twins who grew up in the same home.
Another study, the Texas Adoption Project , followed about 300 children who were adopted within a few days of birth and grew up entirely with their adopted families. John Loehlin and his coworkers at the University of Texas at Austin looked at results of intelligence tests given to the children at age seven and at age 17, to their adoptive parents, and to their biological mothers. The test scores of the biological mothers correlated significantly with those of their children who had been given away for adoption 17 years earlier, whereas there was no correlation between intelligence scores of the children and their adoptive parents. The researchers estimated that 78 percent of intelligence is inherited.
Addictive behavior, or the predilection to become addicted, is another trait whose heritability is controversial. Addiction includes physical dependence and symptoms of craving after chronic substance use, as well as behavioral dependence — the inability to stop an activity even though the consequences are severe. Alcohol, tobacco, and illicit drug use is estimated to contribute to one in eight deaths worldwide.
Yet most people who try habit-forming substances do not become addicted. One study estimates that the probability that someone who tries a substance once will become dependent on it ranges from about one in three or four for tobacco and heroin to about one in six or seven for cocaine and alcohol to about one in 11 for marijuana. The effect of genetics on the vulnerability of individuals to becoming addicted to these substances varies widely. Studies of twins suggest that persistent smoking and nicotine dependence is about 70 percent heritable, alcohol dependence is 50 to 60 percent heritable, and addiction to most other substances is 20 to 35 percent heritable. These studies also indicate that other disorders such as antisocial personality disorder and conduct disorder are often associated with addictive behavior.
Studies of twins suggest that persistent smoking and nicotine dependence is about 70 percent heritable, alcohol dependence is 50 to 60 percent heritable, and addiction to most other substances is 20 to 35 percent heritable.
Psychiatric disorders often have a large — sometimes, surprisingly large — genetic component. Schizophrenia, for which the lifetime risk is approximately 1 percent, has a heritability estimated at around 85 percent. Bipolar disorder (also known as manic-depressive illness because it is characterized by episodes of extreme elation (mania) alternating with episodes of depression) also has an individual lifetime risk of about one percent. Twin, sibling, and family studies all point to a strong genetic basis of bipolar disorder: a heritability of 80 to 90 percent. Depression (also known as unipolar disorder), for which we have an individual lifetime risk of around 10 to 20 percent in the United States, undoubtedly has a genetic basis: Its heritability may be as high as 70 percent.
Susan Middlebrook’s unusual sleep-wake cycle is obviously heritable. This syndrome has revealed something striking about circadian rhythms. Much of what’s known about these rhythms was originally worked out in Drosophila melanogaster, the tiny fruit fly that buzzes about the bananas left on a kitchen counter. Remarkably, the fly’s clockwork mechanism, the proteins it uses to reset its timing each day, works much like ours does. The first mutations to be identified that affect these rhythms, which caused flies to have either a shorter or a longer cycle, or no rhythmic cycle at all, were in a gene given the name period .
In 2001, Louis Ptácek, Ying-Hui Fu, and their colleagues at the University of Utah identified a human mutation that causes FASPS. It turned out to be in a gene that encodes a protein with an amino acid sequence very similar to the fly period protein, so the human gene was dubbed Period2 . This was extraordinary evidence of gene conservation over hundreds of millions of years of evolution. Even stronger evidence came from the finding that the single amino acid change in the Period2 protein that advanced Susan Middlebrook’s clock corresponds to a mutation in fruit flies that advances the clock of that simple organism.
What about the heritability of obsessive-compulsive disorder, the illness that struck Jean and Jill? This disease, affecting about 2 percent of the population, has remarkably diverse symptoms but generally includes four major ones: obsessions and checking behavior; a need for symmetry and order; excessive washing; and hoarding tendencies. Individuals differ in their age of onset, the duration of the illness, and the types of symptoms they display. Many sufferers have tics, and some also have depression, phobias, separation anxiety, or disruptive behavior. Some are troubled by harmful sexual or religious obsessions, or by trichotillomania, compulsive hair pulling. Others may exhibit grooming behaviors.
This diversity of symptoms suggests that obsessive-compulsive disorder may exist in different forms that have different genetic causes — in other words, there may be multiple genetic routes to a group of disorders that have all been given a single name. So it is not surprising that twin studies almost always point to a substantial heritable component for this illness, ranging from 25 percent to 80 percent. Thus, the most probable explanation for Jean and Jill’s similar illnesses at similar ages is the young women’s genetic similarity, although other factors may have contributed to their condition.
Autism is a heart-breaking diagnosis for a parent to receive, and so it was for Lewis’s parents. The prevalence of autism went up more than five-fold in the 1990s, and now may be as high as 1 percent of children. Much of the increase may be due to broader diagnostic criteria and increased physician and parent awareness.
Numerous studies indicate that genetic factors are the main cause of autism. For example, siblings of an autistic child have about 10 times the risk of having the syndrome as the general population. A twin study in the United Kingdom yielded a heritability estimate of more than 90 percent for a broad set of autistic symptoms. Many genes are suspected of being involved in the syndrome. They haven’t been identified yet, but when they are, the diagnosis of autism, which can be difficult and often confusing, will be easier and more precise. Accurate and early diagnosis will also enable earlier intervention, which will greatly improve the prognosis for these children.
Estimating the genetic component of human behaviors and psychiatric disorders may help to remove a lingering stigma attached to people with mental illness — a misplaced sense that these are character flaws — and it may inspire more people to seek treatment. In addition, these studies all point to environmental components — generally still to be teased out — that interact with the genetic ones. That’s good news, since we have some control over the environmental inputs to disease. It is notable that even for the most heritable illnesses, such as schizophrenia or bipolar disorder, the heritability is never 100 percent, and even identical twins are never 100 percent concordant. Thus, even when genetics has a strong effect, it is not absolutely deterministic, so hope should never be abandoned. Finally, identification of the gene variants contributing to a disorder can assist and sharpen diagnosis, which will lead to earlier and possibly more effective treatments.
A high heritability does not imply that just a single gene is involved. In fact, for most of these disorders it is clear that many genes are involved, and no single gene is likely to explain most of the variability. By establishing a genetic basis for these disorders and identifying families that carry the causative changes in their DNA, geneticists can pinpoint the genes that are responsible.
Identification of such genes often leads to insight into the disease that can be tested in organisms suitable for experimentation, such as the fruit fly or the mouse. Analysis of those genes is sure to illuminate disease mechanisms. Just knowing what the relevant genes are will allow individuals to be tested for the gene variants that put them at risk of the disease, and, should they carry some of those variants in their personal DNA code, make them or their parents vigilant about early signs of the disease. And the genes and proteins that are implicated provide potential targets for new drugs that promise to improve the lives of people like Susan, Jean and Jill, Lewis, and many of the rest of us.
Stanley Fields is Professor of Genome Sciences and Medicine at the University of Washington and a Howard Hughes Medical Institute Investigator. Mark Johnston is Professor and Chair of the Department of Biochemistry and Molecular Genetics at the University of Colorado School of Medicine and Editor-in-Chief of the journal Genetics. Fields and Johnston are the authors of “ Genetic Twists of Fate ,” from which this article is excerpted.
| Generation ships offer a tantalizing possibility: transporting humans on a permanent voyage to a new home among the stars. | | |
| A growing body of research suggests that optimism plays a significant role in promoting both physical and mental well-being. | | |
| An excerpt from Emmanuelle Pouydebat’s “Sexus Animalis,” an illustrated guide to the amazingly multifarious sex lives of animals. | | |
| The macabre diets of scale-eating cichlids help shed light on the important role of frequency dependence in shaping genetic variation and the natural world. | |

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
- My Bibliography
- Collections
- Citation manager

Save citation to file
Email citation, add to collections.
- Create a new collection
- Add to an existing collection
Add to My Bibliography
Your saved search, create a file for external citation management software, your rss feed.
- Search in PubMed
- Search in NLM Catalog
- Add to Search
Overview of Behavioral Genetics Research for Family Researchers
Affiliation.
- 1 Department of Psychology, N218 Elliot Hall, 75 East River Road, Minneapolis, MN 55108 ( [email protected] ).
- PMID: 24073018
- PMCID: PMC3780434
- DOI: 10.1111/jftr.12013
This article provides an overview of the methods, assumptions, and key findings of behavioral genetics methodology for family researchers with a limited background. We discuss how family researchers can utilize and contribute to the behavioral genetics field, particularly in terms of conducting research that seeks to explain shared environmental effects. This can be done, in part, by theoretically controlling for genetic confounds in research that seeks to determine cause-and-effect relationships among family variables and individual outcomes. Gene-environment correlation and interaction are especially promising areas for the family researcher to address. Given the methodological advancements in the field, we also briefly comment on new methods in molecular genetics for studying psychological mental health disorders.
Keywords: Behavior genetics; genetic relatedness; human development; shared environment.
PubMed Disclaimer
Figure 1. Diagram of univariate decomposition
Variance components are represented by capital letters (additive genetic:…
Similar articles
- Genetic Influences on Peer and Family Relationships Across Adolescent Development: Introduction to the Special Issue. Mullineaux PY, DiLalla LF. Mullineaux PY, et al. J Youth Adolesc. 2015 Jul;44(7):1347-59. doi: 10.1007/s10964-015-0306-0. Epub 2015 May 26. J Youth Adolesc. 2015. PMID: 26006709
- Research review: the shared environment as a key source of variability in child and adolescent psychopathology. Burt SA. Burt SA. J Child Psychol Psychiatry. 2014 Apr;55(4):304-12. doi: 10.1111/jcpp.12173. Epub 2013 Nov 22. J Child Psychol Psychiatry. 2014. PMID: 24261560 Review.
- Environment, genes, and experience: lessons from behavior genetics. Barsky PI. Barsky PI. J Physiol Paris. 2010 Nov;104(5):243-52. doi: 10.1016/j.jphysparis.2010.08.002. Epub 2010 Sep 8. J Physiol Paris. 2010. PMID: 20816769 Review.
- Genetic and environmental influences on human psychological differences. Bouchard TJ Jr, McGue M. Bouchard TJ Jr, et al. J Neurobiol. 2003 Jan;54(1):4-45. doi: 10.1002/neu.10160. J Neurobiol. 2003. PMID: 12486697 Review.
- The Human Salivary Microbiome Is Shaped by Shared Environment Rather than Genetics: Evidence from a Large Family of Closely Related Individuals. Shaw L, Ribeiro ALR, Levine AP, Pontikos N, Balloux F, Segal AW, Roberts AP, Smith AM. Shaw L, et al. mBio. 2017 Sep 12;8(5):e01237-17. doi: 10.1128/mBio.01237-17. mBio. 2017. PMID: 28900019 Free PMC article.
- Addressing diversity and inclusion challenges in global neuro-psychiatric and behavioral genomics research. Matshabane OP, Whitted CG, Koehly LM. Matshabane OP, et al. Front Genet. 2022 Dec 13;13:1021649. doi: 10.3389/fgene.2022.1021649. eCollection 2022. Front Genet. 2022. PMID: 36583023 Free PMC article.
- Parent involvement, sibling companionship, and adolescent substance use: A longitudinal, genetically informed design. Samek DR, Rueter MA, Keyes MA, McGue M, Iacono WG. Samek DR, et al. J Fam Psychol. 2015 Aug;29(4):614-23. doi: 10.1037/fam0000097. J Fam Psychol. 2015. PMID: 26030026 Free PMC article.
- Homosexual inclinations and the passions: A Thomistic theory of the psychogenesis of same-sex attraction disorder. Kinney RL 3rd. Kinney RL 3rd. Linacre Q. 2014 May;81(2):130-61. doi: 10.1179/2050854914Y.0000000018. Linacre Q. 2014. PMID: 24899749 Free PMC article.
- Agrawal A, Balasubramanian S, Smith EK, Madden PAF, Bucholz KK, Heath AC, Lynskey MT. Peer substance involvement modifies genetic influences on regular substance involvement in young women. Addiction. 2010;105:1844–1853. doi:10.1111/j.1360-0443.2010.02993.x. - PMC - PubMed
- Bazzett TJ. An introduction to behavior genetics. Sinauer; Sunderland, MA: 2008.
- Beaver KM, DeLisi M, Wright JP, Vaughn MG. Gene–environment interplay and delinquent involvement: Evidence of direct, indirect, and interactive effects. Journal of Adolescent Research. 2009;24:147–168. doi:10.1177/0743558408329952.
- Beaver KM, Gibson CL, Turner MG, DeLisi M, Vaughn MG, Holand A. Stability of peer associations: A biosocial test of Warr’s stick-friends hypothesis. Crime and Delinquency. 2009;57:907–926. doi:10.1177/0011128709332660.
- Behavior Genetics Association (n.d.). Retrieved from http://www.bga.org/
Related information
Grants and funding.
- T32 MH017069/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full text sources.
- Europe PubMed Central
- PubMed Central
Other Literature Sources
- scite Smart Citations
- Citation Manager
NCBI Literature Resources
MeSH PMC Bookshelf Disclaimer
The PubMed wordmark and PubMed logo are registered trademarks of the U.S. Department of Health and Human Services (HHS). Unauthorized use of these marks is strictly prohibited.
This page has been archived and is no longer being updated regularly.
Cover Story
A second look at twin studies
As behavioral genetics enters a second century, the field's oldest research method remains both relevant and controversial.
By LEA WINERMAN
Monitor Staff
April 2004, Vol 35, No. 4
Print version: page 46
"Twins have a special claim upon our attention; it is, that their history affords means of distinguishing between the effects of tendencies received at birth, and those that were imposed by the special circumstances of their after lives."
-- Sir Francis Galton, 19th century behavioral genetics pioneer, Inquiries into Human Faculty and its Development, 1875
More than a century after Galton's observation, twin studies remain a favorite tool of behavioral geneticists. Researchers have used twin studies to try to disentangle the environmental and genetic backgrounds of a cornucopia of traits, from aggression to intelligence to schizophrenia to alcohol dependence.
But despite the popularity of twin studies, some psychologists have long questioned assumptions that underlie them--like the supposition that fraternal and identical twins share equal environments or that people choose mates with traits unlike their own. The equal environments assumption, for example, has been debated for at least 40 years. Many researchers have found evidence that the assumption is valid, but others remain skeptical (see Further Reading below).
Overall, twin studies assumptions remain controversial, says psychologist James Jaccard, PhD, a psychologist who studies statistical methods at the University at Albany of the State University of New York. In response, though, researchers are working to expand and develop twin study designs and statistical methods. And while the assumptions question remains a stumbling block for some researchers, many agree twin studies will continue to be an important tool--along with emerging genome and molecular research methods (see article page 42 )--in shedding light on human behavioral genetics.
Methods and theory
The classical twin study design relies on studying twins raised in the same family environments. Monozygotic (identical) twins share all of their genes, while dizygotic (fraternal) twins share only about 50 percent of them. So, if a researcher compares the similarity between sets of identical twins to the similarity between sets of fraternal twins for a particular trait, then any excess likeness between the identical twins should be due to genes rather than environment.
Researchers use this method, and variations on it, to estimate the heritability of traits: The percentage of variance in a population due to genes. Modern twin studies also try to quantify the effect of a person's shared environment (family) and unique environment (the individual events that shape a life) on a trait.
The assumptions those studies rest on--questioned by some psychologists, including, in recent work, Jaccard--include:
Random mating. Twin researchers assume that people are as likely to choose partners who are different from themselves as they are to choose partners who are similar for a particular trait. If, instead, people tend to choose mates like themselves, then fraternal twins could share more than 50 percent of their genes--and hence more similarities on genetically influenced traits--because they would receive similar genes from their mothers and fathers.
Equal environments. Twin researchers also assume that fraternal and identical twins raised in the same homes experience equally similar environments. But some research suggests that parents, teachers, peers and others may treat identical twins more similarly than fraternal twins.
Gene-environment interaction. Some researchers think that interactions between genes and environment, rather than genes and environment separately, may influence many traits. A recent study from Science (Vol. 297, No. 5582) by Avshalom Caspi, PhD, of King's College London, for example, suggests that a gene might moderate propensity for violence, particularly in people who are severely maltreated as children. Many twin study designs don't take this type of complication into account.
Genetic mechanisms. Traits can be inherited through different genetic mechanisms. For traits governed by dominant genetic mechanisms, a dominant gene inherited from one parent trumps a recessive gene inherited from the other parent: If a person inherits a recessive gene for blue eyes from one parent and a dominant gene for brown eyes from the other parent, then the dominant brown gene wins, and the person's eyes are brown.
Additive genetic mechanisms, in contrast, mix together--a plant that receives one red gene and one white gene might, if the genes are additive, turn out pink. Epistatic mechanisms are complex cases where interactions among multiple genes may determine the outcome of one trait. Twin studies, in general, assume that only one type of genetic mechanism--usually additive--is operating for a particular trait.
The value of twin studies
Twin researchers acknowledge that these and other limitations exist. But, they say, the limitations don't negate the usefulness of twin studies. For traits that are substantially influenced by heredity, the approximately two-fold difference in genetic similarity between the two types of twins should outweigh any complications, says John Hewitt, PhD, director of the Institute for Behavioral Genetics at the University of Colorado at Boulder.
And the extent to which different assumptions matter may depend on which trait is being studied. Studies have suggested, for example, that people are more likely to select mates with similar levels of intelligence than they are mates with similar levels of neuroticism, extraversion and other personality traits (see page 50 ). So, researchers who use twins to study intelligence might have to worry more about nonrandom mating than researchers who study personality.
Twin study designs and statistical analysis methods are also constantly evolving and improving. The original twin study design has expanded to include studies of twins' extended families, longitudinal studies and other variations. Some of these variations allow researchers to address previous limitations--they can investigate the effects of nonrandom mating, for example, by including the spouses of twins in studies. In fact, says psychologist Dorret Boomsma, PhD, of Vrije Universiteit in the Netherlands, all of these assumptions can be tested, given the proper data. She argues that they should not be seen as assumptions at all, but instead as mechanisms whose relevance can be tested using study designs that go beyond the classical twin study design.
Analysis methods, likewise, don't remain static. "People are always thinking about ways to improve the analyses," Hewitt says.
Jaccard acknowledges that this is true. "For some designs, we don't have to make as strong assumptions as we used to make," he says. "Instead of having to assume away four constructs, we only have to assume away two or three."
In the age of molecular genetics, meanwhile, the classical twin study design is only one aspect of genetics research. Twin studies estimate the heritability of a trait, but molecular genetics attempts to pinpoint the effects of a particular gene.
The future of twin research will involve combining traditional twin studies with molecular genetics research, according to Hewitt, who believes that day is already here.
"When we conduct a study of twins these days, we always get DNA on everyone," Hewitt says. "And we'll use that DNA to try and identify specific individual genes that contribute to the overall pattern of heritability."
Further Reading
Boomsma, D., Busjahn, A., & Peltonen, L. (2002). Classical twin studies and beyond. Nature Reviews Genetics, 3 (11), 872-882.
Kendler, K.S., Neale, M.C., Kessler, R.C., Heath, A.C., & Eaves, L.J. (1993). A test of the equal-environment assumption in twin studies of psychiatric illness. Behavior Genetics, 23 , 21-28.
Neale, M.C., & Cardon, L.R. (1992). Methodology for genetic studies of twins and families. Dordrecht, The Netherlands: Kluwer Academic Press.
Pam, A., Kemker, S.S., Ross, C.A., & Golden, R. (1996). The "equal environments assumption" in MZ-DZ twin comparisons: An untenable premise of psychiatric genetics? Acta Geneticae Medicae et Gemellologiae, 45 (3), 349-360.
Letters to the Editor
- Send us a letter
Celebrating a Century of Research in Behavioral Genetics
- Open access
- Published: 20 January 2023
- Volume 53 , pages 75–84, ( 2023 )
Cite this article
You have full access to this open access article

- Robert Plomin 1
8159 Accesses
3 Citations
55 Altmetric
Explore all metrics
A century after the first twin and adoption studies of behavior in the 1920s, this review looks back on the journey and celebrates milestones in behavioral genetic research. After a whistle-stop tour of early quantitative genetic research and the parallel journey of molecular genetics, the travelogue focuses on the last fifty years. Just as quantitative genetic discoveries were beginning to slow down in the 1990s, molecular genetics made it possible to assess DNA variation directly. From a rocky start with candidate gene association research, by 2005 the technological advance of DNA microarrays enabled genome-wide association studies, which have successfully identified some of the DNA variants that contribute to the ubiquitous heritability of behavioral traits. The ability to aggregate the effects of thousands of DNA variants in polygenic scores has created a DNA revolution in the behavioral sciences by making it possible to use DNA to predict individual differences in behavior from early in life.
Similar content being viewed by others

Challenges and Strategies for Integrating Molecular Genetics into Behavioral Science
Genotype-environment correlation in the era of dna.

Behavioural genetics methods
Avoid common mistakes on your manuscript.
Introduction
Although the history of heredity and behavior can be traced back to ancient times (Loehlin 2009 ), the first human behavioral genetic research was reported in the 1920s, which applied quantitative genetic twin and adoption designs to assess genetic influence on newly developed measures of intelligence. The 1920s also marked the beginning of single-gene research that led to molecular genetics. The goal of this review is to outline 100 years of progress in quantitative genetic and molecular genetic research on behavior, a whistle-stop tour of a few of the major milestones in the journey. The review focuses on human research even though non-human animal research played a major role in the first 50 years (Maxson 2007 ). It uses intelligence as a focal example because intelligence was the target of much human research, even though a similar story could be told for other areas of behavioral genetics such as psychopathology.
The Two Worlds of Genetics
The most important development during this century of behavioral genetic research has been the synthesis of the two worlds of genetics, quantitative genetics and molecular genetics. Quantitative genetics and molecular genetics both have their origins in the 1860s with Francis Galton (Galton 1865 , 1869 ) and Gregor Mendel (Mendel 1866 ), respectively. Not much happened until the 1900s when Galton’s insights led to methods to study genetic influence on complex traits and when Mendel’s work was re-discovered. The two worlds clashed as Mendelians looked for 3:1 segregation ratios indicative of single-gene traits, whereas Galtonians assumed that Mendel’s laws of heredity were specific to pea plants because they knew that complex traits are distributed continuously.
Antipathy between the two worlds of genetics followed because of the different goals of Mendelians and Galtonians. Mendelians, the predecessors of molecular geneticists, wanted to understand how genes work, which led to the use of induced mutations and a focus on dichotomous traits that were easily assessed such as physical characteristics rather than behavioral traits. In contrast, Galtonians, whose descendants are quantitative geneticists, used genetics as a tool to understand the etiology of naturally occurring variation in complex traits selected for their intrinsic interest and importance, with behavioral traits, especially intelligence, high on the list. The resolution to the conflict could be seen in Ronald Fisher’s 1918 paper, which showed that Mendelian inheritance is compatible with quantitative traits if the assumption is made that several genes affect a trait (Fisher 1918 ). Nonetheless, the two worlds of genetics went their own way for most of the century.
The synthesis of the two worlds of genetics began in the 1980s with the technological advances of DNA sequencing, polymerase chain reaction, and DNA microarrays that enabled genome-wide association (GWA) studies of complex traits. In addition to finding DNA variants associated with complex traits, GWA genotypes led to three far-reaching advances in genetic research. First, GWA genotypes were used to estimate directly the classical quantitative genetic parameters of heritability and genetic correlation, which could be called quantitative genomics . Second, the results of GWA studies were used to create polygenic scores that predict individual differences for complex traits. Third, GWA genotypes facilitated new approaches to causal modeling of the interplay between genes and environment. Together, when applied to behavioral traits, these advances could be called behavioral genomics . This synthesis of the two worlds of genetics, the journey from behavioral genetics to behavioral genomics, is the overarching theme of this whistle-stop tour celebrating a century of research in behavioral genetics. (See Fig. 1 .) The itinerary begins with milestones in quantitative genetics and then molecular genetics, concluding with behavioral genomics.

Synthesis of the two worlds of genetics: from behavioral genetics to behavioral genomics.
Quantitative Genetics
The first 50 years of quantitative genetic research, from 1920 to 1970, started off well with family studies (Jones 1928 ; Thorndike 1928 ), twin studies (Holzinger 1929 ; Lauterbach 1925 ; Merriman 1924 ; Tallman 1928 ) and adoption studies (Burks 1928 ; Freeman et al. 1928 ) using the recently devised IQ test. However, this nascent research was squelched with the emergence of Nazi eugenic policies (McGue 2008 ). The void was filled with behaviorism (Watson 1930 ), which led to environmentalism, the ‘blank slate’ view that we are what we learn (Pinker 2003 ).
Nonetheless, a few studies of IQ appeared in the 1930 and 1940 s, such as the first study of identical twins reared apart (Newman et al. 1937 ) and the first adoption study that assessed birth parents (Skodak and Skeels 1949 ). Both indicated substantial genetic influence on IQ, as did a review of all available IQ data (Woodworth 1941 ).
In 1960, the field-defining book, Behavior Genetics (Fuller and Thompson 1960 ), was published. It mostly reviewed research on nonhuman animals. In their preface, the authors noted that “we considered omitting human studies completely” (p. vi); even their chapter on cognitive abilities primarily reviewed nonhuman research. An earlier influential review began by saying, “In the writer’s opinion, the genetics of behavior must be worked out on species that can be subjected to controlled breeding. At the present time this precludes human subjects” (Hall 1951 ).
In 1963, a milestone review was published in Science of 52 family, twin and adoption studies of IQ (Erlenmeyer-Kimling and Jarvik 1963 ). Although the studies were very small by modern standards and heritability was not calculated, the average results from the different designs suggested substantial heritability. For example, the average MZ and DZ twin correlations were 0.87 and 0.53, respectively, suggesting a heritability of 68%. However, despite being published in Science , the paper was largely ignored; it was cited only 22 times in five years.
The pace of behavioral genetic research picked up in the 1960s, once again primarily research on non-human animals (Lindzey et al. 1971 ; McClearn 1971 ), although some twin studies on cognitive abilities were also published (Nichols 1965 ; Schoenfeldt 1968 ). However, the first 50 years of quantitative genetic research ended badly with the publication in 1969 of Arthur Jensen’s paper, How Much Can We Boost IQ and Scholastic Achievement? (Jensen 1969 ). The paper touched on ethnic differences, which made it one of the most controversial papers in the behavioral sciences, with 900 citations in the first five years and more than 6200 citations in total.
1970 was a watershed year marking the second 50 years of behavioral genetic research. It was the year that the Behavior Genetics Association was launched and the first issue of its journal, Behavior Genetics , was published. Another 1970 milestone was the publication of the foundational paper for model-fitting analysis of quantitative genetic designs (Jinks and Fulker 1970 ).
The 1970s and 1980s yielded most of the major discoveries for quantitative genetics as applied to behavioral traits, discoveries that are listed as landmarks in the following paragraphs. Nonetheless, in the aftermath of Jensen’s 1969 paper, behavioral genetic research, especially on intelligence, was highly controversial (Scarr and Carter-Saltzman 1982 ). Most notably, Leon Kamin severely criticized the politics as well as science of behavioral genetic research on intelligence in his book, The Science and Politics of I.Q. (Kamin 1974 ). He concluded that “There exist no data which should lead a prudent man to accept the hypothesis that I.Q. test scores are in any degree heritable” (p. 1). The book was cited more than 2000 times and stoked antipathy towards genetic research. It also impugned the motivation of genetic researchers, saying that they are ‘committed to the view that those on the bottom are genetically inferior victims of their own immutable defects’ (p. 2).
All Traits are Heritable
Despite this hostility, genetic research grew exponentially in the 1970s and created a seismic shift from the prevailing view that behavioral traits like intelligence are not “in any degree heritable”. In 1978, a review of 30 twin studies of intelligence yielded an average heritability estimate of 46% (Nichols 1978 ). Moreover, the conclusion began to emerge that all traits show substantial heritability. This conclusion, which has been called the first law of behavioral genetics (Turkheimer 2000 ), was first observed in 1976 in a twin study of cognitive data for 3000 twin pairs, which also included extensive data on personality and interests for 850 twin pairs (Loehlin and Nichols 1976 ). The authors noted “the curious uniformity of identical-fraternal differences both within and across trait domains” (p. 89). A 2015 meta-analysis of all published twin studies showed that behavioral traits are about 50% heritable on average (Polderman et al. 2015 ). Demonstrating the ubiquitous importance of genetics was the fundamental accomplishment of behavioral genetics.
No Traits are 100% Heritable
The flip side of the finding of 50% heritability was just as important: no traits are 100% heritable. It is ironic that, after a century of environmentalism, genetic research provided the strongest evidence for the importance of the environment; previous environmental research was confounded because it ignored genetics. Moreover, investigating environmental influences in genetically sensitive designs led to two of the most important discoveries about the environment: nonshared environment and the nature of nurture.
Nonshared Environment
Quantitative genetic research showed that environmental influences work very differently from the way they were assumed to work. A second discovery by Loehlin and Nichols ( 1976 ) was that salient environmental influences are not shared by twins growing up in the same family: “Environment carries substantial weight in determining personality – it appears to account for at least half the variance – but that environment is one for which twin pairs are correlated close to zero” (p. 92). This phenomenon has come to be known as nonshared environment (Plomin and Daniels 1987 ).
Loehlin and Nichols suggested that cognitive abilities are an exception to the rule that environmental influences make children in a family different from, not similar to, one another. Their twin study suggested that about 25% of the variance of cognitive abilities could be attributed to shared environment. A direct test of shared environmental influence is the correlation between adoptive siblings, genetically unrelated children adopted into the same family. Seven small studies of adoptive siblings yielded an average IQ correlation of 0.25, which seemed to precisely confirm the twin estimate (McGue et al. 1993 ).
However, in 1978, a study of 100 pairs of adoptive siblings reported an IQ correlation of -0.03 (Scarr and Weinberg 1978 ). This is a good example of the progressive nature of behavioral genetic research (Urbach 1974 ). Scarr and Weinberg noted that previous studies involved children, whereas theirs was the first study of post-adolescent adoptive siblings aged 16 to 22, and they hypothesized that the effect of shared environmental influence on cognitive development diminishes after adolescence as young adults make their own way in the world. Their hypothesis was confirmed in two additional studies of post-adolescent adoptive siblings that yielded an average IQ correlation of -0.01 (McGue et al. 1993 ). Evidence that shared environmental influence declines after adolescence to negligible levels for cognitive abilities has also emerged from twin studies (Briley and Tucker-Drob 2013 ; Haworth et al. 2010 ). However, one of the biggest mysteries about nonshared environment remains: what are these environmental influences that make children growing up in the same family so different (Plomin 2011 )?
The Nature of Nurture
Another milestone was the revelation that environmental measures widely used in the behavioral sciences, such as parenting, social support, and life events, show genetic influence (Plomin and Bergeman 1991 ), with heritabilities of about 25% on average (Kendler and Baker 2007 ). This finding emerged in the 1980s as measures of the environment were included in quantitative genetic designs, which also led to the discovery that associations between environmental measures and psychological traits are significantly mediated genetically (Plomin et al. 1985 ). The nature of nurture is one of the major directions for research in behavioral genomics, as discussed later.
Heritability Increases During Development
Another milestone in the 1970s was the Louisville Twin Study in which mental development of 500 pairs of twins was assessed longitudinally and showed that the heritability of intelligence increases from infancy to adolescence (Wilson 1983 ). In light of the replication crisis in science (Ritchie 2021 ), a cause for celebration is that this counterintuitive finding of increasing heritability of intelligence – from about 40% in childhood to more than 60% in adulthood -- has consistently replicated, as seen in cross-sectional (Haworth et al. 2010 ) and longitudinal (Briley and Tucker-Drob 2013 ) mega-analyses.
In 1977, a landmark paper showed how univariate analysis of variance can be extended to multivariate analysis of covariance in a model-fitting framework (Martin and Eaves 1977 ). They applied their approach to cognitive abilities and found an average genetic correlation of 0.52, indicating that many genes affect diverse traits, called pleiotropy . Subsequent studies also yielded genetic correlations greater than 0.50 between diverse cognitive abilities (Plomin and Kovas 2005 ).
In the 1970s and 1980s, bigger and better studies made most of the major quantitative genetic discoveries, going far beyond merely estimating heritability. But it was not all smooth sailing. Most notably, The Bell Curve resurrected many of the issues that followed Jensen’s 1969 paper (Herrnstein and Murray 1996 ). Nonetheless, by the 1990s, quantitative genetic research had convinced most scientists of the importance of genetics for behavioral traits, including intelligence (Snyderman and Rothman 1990 ). One symbol of this change was that the 1992 Centennial Conference of the American Psychological Association chose behavioral genetics as one of two themes that best represented the past, present, and future of psychology (Plomin and McClearn 1993 ). Then, just as quantitative genetic discoveries began to slow, the synthesis with molecular genetics began, which led to the DNA revolution and behavioral genomics.
Molecular Genetics
During its first 50 years, molecular genetics focused on single-gene disorders. In 1933, a Nobel prize was awarded to Thomas Hunt Morgan for mapping genes responsible for single-gene mutations in fruit flies (Morgan et al. 1923 ), but human mapping was stymied because only a few single-gene markers such as blood types were available – variants in DNA itself were not available for another fifty years. Research on single-gene effects discovered in pedigree studies only incidentally involved behavioral traits. For example, phenylketonuria, the most common single-gene metabolic disorder, was discovered in 1934 (Folling 1934 ) and shown to be responsible for 1% of the population institutionalized for severe intellectual disability.
In the 1940s, it became clear that DNA is the mechanism of heredity, culminating in the most famous paper in biology which proposed the double-helix structure of DNA (Watson and Crick 1953 ). An important milestone for human behavioral genetics was the discovery in 1959 that the most common form of intellectual disability, Down syndrome, was due to a trisomy of chromosome 21 (Lejeune et al. 1959 ).
In 1961, the genetic code was cracked showing that three-letter sequences of the four-letter alphabet of DNA coded for the 20 amino acids (Crick et al. 1961 ). Just as with quantitative genetics, the 1970s was a watershed decade that ushered in the second 50 years, the genomics era.
The Genomics Era
The era of genomics began in the 1970s when methods were developed to sequence DNA’s nucleotide bases (Sanger et al. 1977 ). In 2003, fifty years after the discovery of the double helix structure of DNA, the Human Genome Project identified the sequence of 92% of the three billion nucleotide bases in the human genome (Collins et al. 2003 ).
In the 1980s, the first common variants in DNA itself were discovered, restriction fragment length polymorphisms (RFLPs) (Botstein et al. 1980 ). RFLPs enabled linkage mapping for single-gene disorders and were the basis for DNA fingerprinting, which revolutionized forensics (Jeffreys 1987 ). Polymerase chain reaction (PCR) was also developed which facilitated genotyping by rapidly amplifying DNA fragments (Mullis et al. 1986 ). In the 1980s, these developments increased the pace of linkage mapping of single-gene disorders, many of which had cognitive consequences, such as phenylketonuria (Woo et al. 1983 ) and Huntington disease (Gusella et al. 1983 ). In the 1990s, DNA sequencing revealed thousands of single-nucleotide polymorphisms (SNPs), the most common DNA variant (Collins et al. 1997 ).
In the 1990s, linkage was also attempted for complex traits that did not show single-gene patterns of transmission, such as reading disability (Cardon et al. 1994 ), but these were unsuccessful because linkage, which traces chromosomal recombination between disease genes and DNA variants within families, is unable to detect small effect sizes (Plomin et al. 1994 ). Researchers then pivoted towards allelic association in unrelated individuals, which is much more powerful in detecting DNA variants of small effect size. An early example of association was an allele of the apolipoprotein E gene on chromosome 19 that was found in 40% of individuals with late-onset Alzheimer disease as compared to 15% in controls (Corder et al. 1993 ).
The downside of allelic association is that an association can only be detected if a DNA variant is itself the functional gene or very close to it. For this reason, and because genotyping each DNA variant was slow and expensive, the 1990s became the decade of candidate gene studies in which thousands of studies reported associations between complex behavioral traits and a few ‘candidate’ genes, typically neurotransmitter genes thought to be involved in behavioral pathways. However, these candidate-gene associations failed to replicate because these studies committed most of the sins responsible for the replication crisis (Ioannidis 2005 ). For example, when 12 candidate genes reported to be associated with intelligence were tested in three large samples, none replicated (Chabris et al. 2012 ).
Genome-wide Association
In 1996, an idea emerged that was the opposite of the candidate-gene approach: using thousands of DNA variants to systematically assess associations across the genome in large samples of unrelated individuals (Risch and Merikangas 1996 ). However, genome-wide association (GWA) seemed a dream because genotyping was slow and expensive.
The problem of genotyping each DNA variant in large samples was solved in the 2000s by the commercial availability of DNA microarrays, called SNP chips , which genotype hundreds of thousands of SNPs for an individual quickly, accurately, and inexpensively. SNP chips paved the way for GWA analyses. In 2007, the first major GWA analysis included 2000 cases for each of seven major disorders and compared SNP allele frequencies for these cases with controls (The Wellcome Trust Case Control Consortium 2007 ). Replicable associations were found but they were few in number and extremely small in effect size. Hundreds of GWA reports appeared in the next decade with similarly small effect sizes across the behavioral and biological sciences (Visscher et al. 2017 ), including cognitive traits such as educational attainment (Rietveld et al. 2013 ) and intelligence in childhood (Benyamin et al. 2014 ) and adulthood (Davies et al. 2011 ).
These GWA studies led to the realization that the biggest effect sizes were much smaller than anyone anticipated. For case-control studies, risk ratios were less than 1.1, and for dimensional traits, variance explained was less than 0.001. This meant that huge sample sizes would be needed to detect these miniscule effects, and thousands of these associations would be needed to account for heritability, which is usually greater than 50% for cognitive traits. Ever larger GWA samples scooped up more of these tiny effects. Most recently, a GWA meta-analysis with a sample size of 3 million netted nearly four thousand independent significant associations after correction for multiple testing, but the median effect size of these SNPs accounted for less than 0.0001 of the variance (Okbay et al. 2022 ).
A century after Fisher’s 1918 paper, the discovery of such extreme polygenicity (Boyle et al. 2017 ; Visscher et al. 2021 ) was a turning point in the voyage from behavioral genetics to behavioral genomics. GWA genotypes brought the two worlds of genetics together by making it possible to use GWA genotypes to create three sets of tools to investigate highly polygenic traits: quantitative genomics, polygenic scores, and causal modeling (see Fig. 1 ). When applied to behavioral traits, these tools constitute the new field of behavioral genomics.
Quantitative Genomics
What good are SNP associations that account for such tiny effects? The molecular genetic goal of tracking effects from genes to brain to behavior is daunting when the effects are so small. However, in contrast to this bottom-up approach from genes to behavior, the top-down perspective of behavioral genetics answered this question by using GWA genotypes to estimate quantitative genetic parameters of heritability and genetic correlations, which could be called quantitative genomics . The journey picked up speed as quantitative genomics led to three new milestones.
Genome-wide Complex Trait Analysis (GCTA). In 2011, the first new method was devised to estimate heritability and genetic correlations since twin and adoption designs in the early 1900s. GCTA (originally called GREML) uses GWA genotypes for large samples of unrelated individuals to compare overall SNP similarity to phenotypic similarity pair by pair for all pairs of individuals (Yang et al. 2011 ). The extent to which SNP similarity explains trait similarity is called SNP heritability because it is limited to heritability estimated by the SNPs on the SNP chip. Genetic correlations are estimated by comparing each pair’s SNP similarity to their cross-trait phenotypic similarity.
SNP heritability estimates are about half the heritability estimated by twin studies (Plomin and von Stumm 2018 ). This ‘missing heritability’ occurs because SNP heritability is limited to the common SNPs genotyped on current SNP chips, which also creates a ceiling for discovery in GWA research. Most SNPs are not common, and rare SNPs appear to be responsible for much of the missing heritability, at least for height (Wainschtein et al. 2022 ). Importantly, quantitative genomic estimates of genetic correlations are not limited in this way and thus provide estimates of genetic correlations similar to those from twin studies (Trzaskowski et al. 2013 ).
Linkage Disequilibrium Score (LDSC) Regression. In 2015, a second quantitative genomic method, LDSC, was published which estimates heritability and genetic correlations from GWA summary effect size statistics for each SNP, corrected for linkage disequilibrium between SNPs (Bulik-Sullivan et al. 2015 ). LDSC estimates of heritability and genetic correlations are similar to GCTA estimates, although GCTA estimates are generally more accurate (Evans et al. 2018 ; Ni et al. 2018 ). The advantage of LDSC is that it can be applied to published GWA summary statistics in contrast to GCTA which requires access to GWA data for individuals in the GWA study.
Genomic Structural Equation Modeling (Genomic SEM). In 2019, a third quantitative genomic analysis completed the arc from quantitative genetics to quantitative genomics by combining quantitative genetic structural equation model-fitting, routinely used in twin analyses, to LDSC heritabilities and genetic correlations (Grotzinger et al. 2019 ). Genomic SEM provides insights into the multivariate genetic architecture of cognitive traits (Grotzinger et al. 2019 ) and psychopathology (Grotzinger et al. 2022 ).
The second answer to the question about what to do with SNP associations that have such small effect sizes is the creation of polygenic scores.
Polygenic Scores
A milestone that marks the spot where the DNA revolution began to transform the behavioral sciences is polygenic scores. Rather than using GWA genotypes to estimate SNP heritabilities and genetic correlations, polygenic scores use GWA genotypes to create a single score for each individual that aggregates, across all SNPs on a SNP chip, an individual’s genotype for each SNP (0, 1 or 2) weighted by the SNP’s effect size on the target trait as indicated by GWA summary statistics. In 2001, polygenic scores were introduced in plant and animal breeding (Meuwissen et al. 2001 ) and later in cognitive abilities (Harlaar et al. 2005 ) and psychopathology (Purcell et al. 2009 ). GWA summary statistics needed to create polygenic scores are now publicly available for more than 500 traits, including dozens for psychiatric disorders and other behavioral traits including cognitive traits (PGS Catalog 2022 ).
The most predictive polygenic scores in the behavioral sciences are for cognitive traits, especially educational attainment and intelligence. Early GWA studies of cognitive traits were underpowered to detect the small effects that we now know are responsible for heritability (Plomin and von Stumm 2018 ). In 2013, a landmark was a GWA study of educational attainment with a sample size exceeding 100,000 (Rietveld et al. 2013 ). A polygenic score derived from its GWA summary statistics predicted 2% of the variance of educational attainment in independent samples. The finding that the biggest effects accounted for only 0.0002 of the variance of educational attainment made it clear that much larger samples would be needed to scoop up more of the tiny effects responsible for the twin heritability estimate of about 40%. In the past decade, the predictive power of polygenic scores for educational attainment has increased with increasing sample sizes from 2% (Rietveld et al. 2013 ) to 5% (Okbay et al. 2016 ) to 10% (Lee et al. 2018 ) to 14% in a GWA study with a sample size of three million (Okbay et al. 2022 ). The current polygenic score for intelligence, derived from a GWA study with a sample of 280,000, predicted 4% of the variance (Savage et al. 2018 ), but, together, the polygenic scores for educational attainment and intelligence predicted 10% of the variance of intelligence test scores (Allegrini et al. 2019 ).
The next milestone will be to narrow the gap between heritability explained by polygenic scores and SNP heritability. A more daunting challenge will be to break through the ceiling of SNP heritability to reach the heritability estimated by twin studies. Reaching both of these destinations will be facilitated by even larger GWA studies and whole-genome sequencing (Wainschtein et al. 2022 ).
Polygenic scores are unique predictors because inherited DNA variations do not change systematically during life – there is no backward causation in the sense that nothing in the brain, behavior or environment changes inherited differences in DNA sequence. For this reason, polygenic scores can predict behavioral traits from early in life without knowing anything about the intervening pathways between genes, brain, and behavior.
Polygenic scores have brought behavioral genetics to the forefront of research in many areas of the life sciences because polygenic scores can be created in any sample of unrelated individuals for whom GWA genotype data are available. No special samples of twins or adoptees are needed, nor is it necessary to assess behavioral traits in order to use polygenic scores to predict them.
Although the implications and applications of polygenic scores derive from its power to predict behavioral traits without regard to explanation (Plomin and von Stumm 2022 ), another milestone on the road to behavioral genomics has been the leverage provided by GWA genotypes for causal modeling.
Causal Modeling
A final milestone on the journey from behavioral genetics to behavioral genomics is a suite of new approaches that use GWA genotypes in causal models that attempt to dissect sources of genetic influence on behavioral traits (Pingault et al. 2018 ). Although traditional quantitative genetic models are causal models, GWA genotypes have enhanced causal modeling in research on assortative mating (Border et al. 2021 ; Yengo et al. 2018 ), population stratification (Abdellaoui et al. 2022 ; Lawson et al. 2020 ), and Mendelian randomization (Richmond and Davey Smith 2022 ).
An explosion of research on genotype-environment correlation was ignited by a 2018 paper in Science on the topic of the nature of nurture (Kong et al. 2018 ). The study included both parent and offspring GWA genotypes and showed that a polygenic score computed from non-transmitted alleles from parent to offspring influenced offspring educational attainment; these indirect effects were dubbed genetic nurture . GCTA has also been used to investigate genotype-environment correlation (Eilertsen et al. 2021 ). Although a great strength of behavioral genomics is its ability to investigate genetic influence in samples of unrelated individuals, combining GWA genotypes with traditional quantitative genetic designs has also enriched causal modeling (McAdams et al. 2022 ), for example, by comparing results within and between families (Brumpton et al. 2020 ; Howe et al. 2022 ).
This whistle-stop tour has highlighted some of the milestones in a century of research in behavioral genetics. The progress is unmatched in the behavioral sciences and its discoveries have been transformative. The most exciting development is the synthesis of quantitative genetics and molecular genetics into behavioral genomics. The energy from this fusion will propel the field far into the future.
Abdellaoui A, Dolan CV, Verweij KJH, Nivard MG (2022) Gene–environment correlations across geographic regions affect genome-wide association studies. Nat Genet 54:1345–1354. https://doi.org/10.1038/s41588-022-01158-0
Article PubMed PubMed Central Google Scholar
Allegrini AG, Selzam S, Rimfeld K et al (2019) Genomic prediction of cognitive traits in childhood and adolescence. Mol Psychiatry 24:819–827. https://doi.org/10.1038/s41380-019-0394-4
Benyamin B, Pourcain Bs, Davis OS et al (2014) Childhood intelligence is heritable, highly polygenic and associated with FNBP1L. Mol Psychiatry 19:253–258. https://doi.org/10.1038/mp.2012.184
Article PubMed Google Scholar
Border R, O’Rourke S, de Candia T et al (2021) Assortative mating biases marker-based heritability estimators. Nat Genet 13:660. https://doi.org/10.1038/s41467-022-28294-9
Botstein D, White RL, Skolnick M, Davis RW (1980) Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am J Hum Genet 32:314–331
PubMed PubMed Central Google Scholar
Boyle EA, Li YI, Pritchard JK (2017) An expanded view of complex traits: from polygenic to omnigenic. Cell 169:1177–1186. https://doi.org/10.1016/j.cell.2017.05.038
Briley DA, Tucker-Drob EM (2013) Explaining the increasing heritability of cognitive ability across development: a meta-analysis of longitudinal twin and adoption studies. Psychol Sci 24:1704–1713. https://doi.org/10.1177/0956797613478618
Brumpton B, Sanderson E, Heilbron K et al (2020) Avoiding dynastic, assortative mating, and population stratification biases in Mendelian randomization through within-family analyses. Nat Commun 11:3519. https://doi.org/10.1038/s41467-020-17117-4
Bulik-Sullivan B, ReproGen C, Psychiatric Genomics Consortium et al (2015) An atlas of genetic correlations across human diseases and traits. Nat Genet 47:1236–1241. https://doi.org/10.1038/ng.3406
Burks BS (1928) The relative influence of nature and nurture upon mental development; a comparative study of foster parent-foster child resemblance and true parent-true child resemblance. Teachers Coll Record 29:219–316. https://doi.org/10.1177/016146812802900917
Article Google Scholar
Cardon LR, Smith SD, Fulker DW et al (1994) Quantitative trait locus for reading disability on chromosome 6. Science 266:276–279. https://doi.org/10.1126/science.7939663
Chabris CF, Hebert BM, Benjamin DJ et al (2012) Most reported genetic associations with general intelligence are probably false positives. Psychol Sci 23:1314–1323. https://doi.org/10.1177/0956797611435528
Collins FS, Green ED, Guttmacher AE, Guyer MS (2003) A vision for the future of genomics research. Nature 422:835–847. https://doi.org/10.1038/nature01626
Collins FS, Guyer MS, Chakravarti A (1997) Variations on a theme: cataloging human dna sequence variation. Science 278:1580–1581. https://doi.org/10.1126/science.278.5343.1580
Corder EH, Saunders AM, Strittmatter WJ et al (1993) Gene dose of apolipoprotein e type 4 allele and the risk of alzheimer’s disease in late onset families. Science 261:921–923. https://doi.org/10.1126/science.8346443
Crick FHC, Barnett L, Brenner S, Watts-Tobin RJ (1961) General nature of the genetic code for proteins. Nature 192:1227–1232. https://doi.org/10.1038/1921227a0
Davies G, Tenesa A, Payton A et al (2011) Genome-wide association studies establish that human intelligence is highly heritable and polygenic. Mol Psychiatry 16:996–1005. https://doi.org/10.1038/mp.2011.85
Eilertsen EM, Jami ES, McAdams TA et al (2021) Direct and indirect effects of maternal, paternal, and offspring genotypes: Trio-GCTA. Behav Genet 51:154–161. https://doi.org/10.1007/s10519-020-10036-6
Erlenmeyer-Kimling L, Jarvik LF (1963) Genetics and intelligence: a review. Science 142:1477–1479. https://doi.org/10.1126/science.142.3598.1477
Evans LM, Tahmasbi R, Vrieze SI et al (2018) Comparison of methods that use whole genome data to estimate the heritability and genetic architecture of complex traits. Nat Genet 50:737–745. https://doi.org/10.1038/s41588-018-0108-x
Fisher RA (1918) The correlation between relatives on the supposition of mendelian inheritance. Trans R Soc Edinb 52:399–433. https://doi.org/10.1017/S0080456800012163
Folling A (1934) Excretion of phenylpyruvic acid in urine as a metabolicnomaly in connection with imbecility.Nord Med Tidskr 8:1054-1059
Freeman FN, Holzinger KJ, Mithell BC (1928) The influence of environment on the intelligence, school achievement, and conduct of foster children. Yearbook of the National Society for the Study of Education 1:102–217
Google Scholar
Fuller JL, Thompson WR (1960) Behavior genetics. Wiley, New York
Book Google Scholar
Galton F (1865) Hereditary talent and character. Macmillan’s Magazine 12:157–166
Galton F (1869) Hereditary genius: an inquiry into its laws and consequences. Collins, London
Grotzinger AD, Mallard TT, Akingbuwa WA et al (2022) Genetic architecture of 11 major psychiatric disorders at biobehavioral, functional genomic and molecular genetic levels of analysis. Nat Genet 54:548–559. https://doi.org/10.1038/s41588-022-01057-4
Grotzinger AD, Rhemtulla M, de Vlaming R et al (2019) Genomic structural equation modelling provides insights into the multivariate genetic architecture of complex traits. Nat Hum Behav 3:513–525. https://doi.org/10.1038/s41562-019-0566-x
Gusella JF, Wexler NS, Conneally PM et al (1983) A polymorphic DNA marker genetically linked to Huntington’s disease. Nature 306:234–238. https://doi.org/10.1038/306234a0
Hall CS (1951) The genetics of behavior. In: Stevens SS (ed) Handbook of experimental psychology. Wiley, New York, pp 304–329
Harlaar N, Butcher LM, Meaburn E et al (2005) A behavioural genomic analysis of DNA markers associated with general cognitive ability in 7-year-olds. J Child Psychol & Psychiat 46:1097–1107. https://doi.org/10.1111/j.1469-7610.2005.01515.x
Haworth CMA, Wright MJ, Luciano M et al (2010) The heritability of general cognitive ability increases linearly from childhood to young adulthood. Mol Psychiatry 15:1112–1120. https://doi.org/10.1038/mp.2009.55
Herrnstein RJ, Murray CA (1996) The bell curve: intelligence and class structure in American life, 1st Free Press pbk. ed. Simon & Schuster, New York
Holzinger KJ (1929) The relative effect of nature and nurture influences on twin differences. J Educ Psychol 20:241–248. https://doi.org/10.1037/h0072484
Howe LJ, Nivard MG, Morris TT et al (2022) Within-sibship genome-wide association analyses decrease bias in estimates of direct genetic effects. Nat Genet 54:581–592. https://doi.org/10.1038/s41588-022-01062-7
Ioannidis JPA (2005) Why most published research findings are false. PLoS Med 2:e124. https://doi.org/10.1371/journal.pmed.0020124
Jeffreys AJ (1987) Highly variable minisatellites and DNA fingerprints. Biochem Soc Trans 15:309–317. https://doi.org/10.1042/bst0150309
Jensen A (1969) How much can we boost IQ and scholastic achievement? Harv Educational Rev 39:1–123. https://doi.org/10.17763/haer.39.1.l3u15956627424k7
Jinks JL, Fulker DW (1970) Comparison of the biometrical genetical, MAVA, and classical approaches to the analysis of the human behavior. Psychol Bull 73:311–349. https://doi.org/10.1037/h0029135
Jones HE (1928) A first study of parent-child resemblance in intelligence. Yearbook of the National Society for the Study of Education Part 1:61–72
Kamin LJ (1974) The science and politics of I.Q. Routledge, New York, NY
Kendler KS, Baker JH (2007) Genetic influences on measures of the environment: a systematic review. Psychol Med 37:615–626. https://doi.org/10.1017/S0033291706009524
Kong A, Thorleifsson G, Frigge ML et al (2018) The nature of nurture: Effects of parental genotypes. Science 359:424–428. https://doi.org/10.1126/science.aan6877
Lauterbach CE (1925) Studies in twin resemblance. Genetics 10:525–568. https://doi.org/10.1093/genetics/10.6.525
Lawson DJ, Davies NM, Haworth S et al (2020) Is population structure in the genetic biobank era irrelevant, a challenge, or an opportunity? Hum Genet 139:23–41. https://doi.org/10.1007/s00439-019-02014-8
Lee JJ, Wedow R, Okbay A et al (2018) Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat Genet 50:1112–1121. https://doi.org/10.1038/s41588-018-0147-3
Lejeune J, Gautier M, Turpin R (1959) [Study of somatic chromosomes from 9 mongoloid children]. C R Hebd Seances Acad Sci 248:1721–1722
PubMed Google Scholar
Lindzey G, Loehlin J, Manosevitz M, Thiessen D (1971) Behavioral genetics. Annu Rev Psychol 22:39–94. https://doi.org/10.1146/annurev.ps.22.020171.000351
Loehlin JC (2009) History of behavior genetics. In: Kim Y-K (ed) Handbook of behavior genetics. Springer New York, New York, NY, pp 3–11
Chapter Google Scholar
Loehlin JC, Nichols RC (1976) Heredity, environment, and personality: a study of 850 sets of twins. University of Texas Press, Austin, Texas
Martin NG, Eaves LJ (1977) The genetical analysis of covariance structure. Heredity 38:79–95. https://doi.org/10.1038/hdy.1977.9
Maxson SC (2007) A history of behavior genetics. In: Jones BC, Mormee P (eds) Neurobehavioral genetics: methods and applications, 2nd edn. CRC Press, New York, pp 1–16
McAdams TA, Cheesman R, Ahmadzadeh YI (2022) Annual Research Review: towards a deeper understanding of nature and nurture: combining family-based quasi‐experimental methods with genomic data. Child Psychol Psychiatry jcpp 13720. https://doi.org/10.1111/jcpp.13720
McClearn GE (1971) Behavioral genetics. Syst Res 16:64–81. https://doi.org/10.1002/bs.3830160106
McGue M (2008) The end of behavioral genetics? Acta Physiol Sinica 40:1073–1087. https://doi.org/10.3724/sp.j.1041.2008.01073
McGue M, Bouchard TJ, Iacono WG, Lykken (1993) Behavioral genetics of cognitive ability: a life-span perspective. In: Plomin R, McClearn GE, McGuffin P (eds) Nature, nurture and psychology. American Psychological Association, Washington, DC, pp 59–76
Mendel G (1866) Versuche ueber Pflanzenhybriden. Verhandllungen des Naturforschhunden Vereines in Bruenn 4:3–47
Merriman C (1924) The intellectual resemblance of twins. Psychol Monogr 33:i–57. https://doi.org/10.1037/h0093212
Meuwissen THE, Hayes BJ, Goddard ME (2001) Prediction of total genetic value using genome-wide dense marker maps. Genetics 157:1819–1829. https://doi.org/10.1093/genetics/157.4.1819
Morgan TH, Sturtevant AH, Muller HJ, Bridges CB (1923) The mechanism of Mendelian heredity. H. Holt and Company
Mullis K, Faloona F, Scharf S et al (1986) Specific enzymatic amplification of DNA in vitro: the polymerase chain reaction. Cold Spring Harb Symp Quant Biol 51:263–273. https://doi.org/10.1101/SQB.1986.051.01.032
Newman HH, Freeman FN, Holzinger KJ (1937) Twins: a study of heredity and environment. University of Chicago Press
Ni G, Moser G, Wray NR et al (2018) Estimation of genetic correlation via linkage disequilibrium score regression and genomic restricted maximum likelihood. Am J Hum Genet 102:1185–1194. https://doi.org/10.1016/j.ajhg.2018.03.021
Nichols RC (1965) The National Merit Twin Study. In: Vandenberg SG (ed) Methods and goals of human behavior genetics. Academic Press, New York, pp 231–243
Nichols RC (1978) Twin studies of ability, personality, and interests. Homo 29:158–173
Okbay A, Beauchamp JP, Fontana MA et al (2016) Genome-wide association study identifies 74 loci associated with educational attainment. Nature 533:539–542. https://doi.org/10.1038/nature17671
Okbay A, Wu Y, Wang N et al (2022) Polygenic prediction of educational attainment within and between families from genome-wide association analyses in 3 million individuals. Nat Genet 54:437–449. https://doi.org/10.1038/s41588-022-01016-z
PGS Catalog (2022) PGS Catalog. http://www.pgscatalog.org/
Pingault J-B, O’Reilly PF, Schoeler T et al (2018) Using genetic data to strengthen causal inference in observational research. Nat Rev Genet 19:566–580. https://doi.org/10.1038/s41576-018-0020-3
Pinker S (2003) The blank slate: the modern denial of human nature. Penguin, London
Plomin R (2011) Commentary: why are children in the same family so different? Non-shared environment three decades later. Int J Epidemiol 40:582–592. https://doi.org/10.1093/ije/dyq144
Plomin R, Bergeman CS (1991) The nature of nurture: genetic influence on “environmental” measures. Behav Brain Sci 14:373–386. https://doi.org/10.1017/S0140525X00070278
Plomin R, Daniels D (1987) Why are children in the same family so different from one another? Behav Brain Sci 10:1–16. https://doi.org/10.1017/S0140525X00055941
Plomin R, Kovas Y (2005) Generalist genes and learning disabilities. Psychol Bull 131:592–617. https://doi.org/10.1037/0033-2909.131.4.592
Plomin R, Loehlin JC, DeFries JC (1985) Genetic and environmental components of “environmental” influences. Dev Psychol 21:391–402. https://doi.org/10.1037/0012-1649.21.3.391
Plomin R, McClearn GE (eds) (1993) Nature, nurture, & psychology. American Psychological Association, Washington, DC
Plomin R, Owen M, McGuffin P (1994) The genetic basis of complex human behaviors. Science 264:1733–1739. https://doi.org/10.1126/science.8209254
Plomin R, von Stumm S (2018) The new genetics of intelligence. Nat Rev Genet 19:148–159. https://doi.org/10.1038/nrg.2017.104
Plomin R, von Stumm S (2022) Polygenic scores: prediction versus explanation. Mol Psychiatry 27:49–52. https://doi.org/10.1038/s41380-021-01348-y
Polderman TJC, Benyamin B, de Leeuw CA et al (2015) Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nat Genet 47:702–709. https://doi.org/10.1038/ng.3285
Purcell SM, Wray NR, Stone JL et al (2009) Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 460:748–752. https://doi.org/10.1038/nature08185
Richmond RC, Davey Smith G (2022) Mendelian randomization: concepts and scope. Cold Spring Harb Perspect Med 12:a040501. https://doi.org/10.1101/cshperspect.a040501
Rietveld CA, Medland SE, Derringer J et al (2013) GWAS of 126,559 individuals identifies genetic variants associated with educational attainment. Science 340:1467–1471. https://doi.org/10.1126/science.1235488
Risch N, Merikangas K (1996) The future of genetic studies of complex human diseases. Science 273:1516–1517. https://doi.org/10.1126/science.273.5281.1516
Ritchie S (2021) Science fictions: exposing fraud, bias, negligence and hype in science. Vintage, London
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA 74:5463–5467. https://doi.org/10.1073/pnas.74.12.5463
Savage JE, Jansen PR, Stringer S et al (2018) Genome-wide association meta-analysis in 269,867 individuals identifies new genetic and functional links to intelligence. Nat Genet 50:912–919. https://doi.org/10.1038/s41588-018-0152-6
Scarr S, Carter-Saltzman L (1982) Genetics and intelligence. In: Sternberg RJ (ed) Handbook of human intelligence. Cambridge University Press, Cambridge, pp 792–895
Scarr S, Weinberg RA (1978) The influence of “family background” on intellectual attainment. Am Sociol Rev 43:674. https://doi.org/10.2307/2094543
Schoenfeldt LF (1968) The hereditary components of the Project TALENT two-day test battery. Meas Evaluation Guidance 1:130–140. https://doi.org/10.1080/00256307.1968.12022379
Skodak M, Skeels HM (1949) A final follow-up study of one hundred adopted children. The Pedagogical Seminary and Journal of Genetic Psychology 75:85–125. https://doi.org/10.1080/08856559.1949.10533511
Snyderman M, Rothman S (1990) The IQ controversy, the media and public policy. Transaction, New Brunswick, NJ
Tallman GG (1928) A comparative study of identical and non-identical twins with respect to intelligence resemblances. Teachers Coll Record 29:83–86. https://doi.org/10.1177/016146812802900912
The Wellcome Trust Case Control Consortium (2007) Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature 447:661–678. https://doi.org/10.1038/nature05911
Article PubMed Central Google Scholar
Thorndike EL (1928) The resemblance of siblings in intelligence. Teachers Coll Record 29:41–53. https://doi.org/10.1177/016146812802900904
Trzaskowski M, Davis OSP, DeFries JC et al (2013) DNA evidence for strong genome-wide pleiotropy of cognitive and learning abilities. Behav Genet 43:267–273. https://doi.org/10.1007/s10519-013-9594-x
Turkheimer E (2000) Three laws of behavior genetics and what they mean. Curr Dir Psychol Sci 9:160–164. https://doi.org/10.1111/1467-8721.00084
Urbach P (1974) Progress and degeneration in the “IQ debate” I and II. Br J Philosophical Sci 25:99–135
Visscher PM, Wray NR, Zhang Q et al (2017) 10 years of GWAS discovery: biology, function, and translation. Am J Hum Genet 101:5–22. https://doi.org/10.1016/j.ajhg.2017.06.005
Visscher PM, Yengo L, Cox NJ, Wray NR (2021) Discovery and implications of polygenicity of common diseases. Science 373:1468–1473. https://doi.org/10.1126/science.abi8206
Wainschtein P, Jain D, Zheng Z et al (2022) Assessing the contribution of rare variants to complex trait heritability from whole-genome sequence data. Nat Genet 54:263–273. https://doi.org/10.1038/s41588-021-00997-7
Watson JB (1930) Behaviorism, revised. University of Chicago Press, Chicago
Watson JD, Crick FHC (1953) Genetical implications of the structure of deoxyribonucleic acid. Nature 171:964–967. https://doi.org/10.1038/171964b0
Wilson RS (1983) The Louisville Twin Study: developmental synchronies in behavior. Child Dev 54:298. https://doi.org/10.2307/1129693
Woo SLC, Lidsky AS, Güttler F et al (1983) Cloned human phenylalanine hydroxylase gene allows prenatal diagnosis and carrier detection of classical phenylketonuria. Nature 306:151–155. https://doi.org/10.1038/306151a0
Woodworth RS (1941) Heredity and environment: a critical survey of recently published material on twins and foster children. Social Science Research Council, New York
Yang J, Manolio TA, Pasquale LR et al (2011) Genome partitioning of genetic variation for complex traits using common SNPs. Nat Genet 43:519–525. https://doi.org/10.1038/ng.823
Yengo L, Robinson MR, Keller MC et al (2018) Imprint of assortative mating on the human genome. Nat Hum Behav 2:948–954. https://doi.org/10.1038/s41562-018-0476-3
Download references
Acknowledgements
This work was supported in part by the UK Medical Research Council (MR/V012878/1 and previously MR/M021475/1).
Author information
Authors and affiliations.
Institute of Psychiatry, Psychology and Neuroscience, King’s College London, London, UK
Robert Plomin
You can also search for this author in PubMed Google Scholar
Contributions
The author wrote, reviewed, and submitted the manuscript.
Corresponding author
Correspondence to Robert Plomin .
Ethics declarations
Conflict of interest.
The author has no relevant financial or non-financial interests to disclose.
Additional information
Publisher’s note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This review was based on a talk given at the 52nd Behavior Genetics Association Annual Meeting, Los Angeles, California, June 25, 2022.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ .
Reprints and permissions
About this article
Plomin, R. Celebrating a Century of Research in Behavioral Genetics. Behav Genet 53 , 75–84 (2023). https://doi.org/10.1007/s10519-023-10132-3
Download citation
Received : 28 December 2022
Revised : 28 December 2022
Accepted : 04 January 2023
Published : 20 January 2023
Issue Date : March 2023
DOI : https://doi.org/10.1007/s10519-023-10132-3
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Quantitative genetics
- Molecular genetics
- Genome-wide association
- Polygenic scores
- Intelligence
- Find a journal
- Publish with us
- Track your research
Information
- Author Services
Initiatives
You are accessing a machine-readable page. In order to be human-readable, please install an RSS reader.
All articles published by MDPI are made immediately available worldwide under an open access license. No special permission is required to reuse all or part of the article published by MDPI, including figures and tables. For articles published under an open access Creative Common CC BY license, any part of the article may be reused without permission provided that the original article is clearly cited. For more information, please refer to https://www.mdpi.com/openaccess .
Feature papers represent the most advanced research with significant potential for high impact in the field. A Feature Paper should be a substantial original Article that involves several techniques or approaches, provides an outlook for future research directions and describes possible research applications.
Feature papers are submitted upon individual invitation or recommendation by the scientific editors and must receive positive feedback from the reviewers.
Editor’s Choice articles are based on recommendations by the scientific editors of MDPI journals from around the world. Editors select a small number of articles recently published in the journal that they believe will be particularly interesting to readers, or important in the respective research area. The aim is to provide a snapshot of some of the most exciting work published in the various research areas of the journal.
Original Submission Date Received: .
- Active Journals
- Find a Journal
- Proceedings Series
- For Authors
- For Reviewers
- For Editors
- For Librarians
- For Publishers
- For Societies
- For Conference Organizers
- Open Access Policy
- Institutional Open Access Program
- Special Issues Guidelines
- Editorial Process
- Research and Publication Ethics
- Article Processing Charges
- Testimonials
- Preprints.org
- SciProfiles
- Encyclopedia
Article Menu

- Subscribe SciFeed
- Recommended Articles
- Google Scholar
- on Google Scholar
- Table of Contents
Find support for a specific problem in the support section of our website.
Please let us know what you think of our products and services.
Visit our dedicated information section to learn more about MDPI.
JSmol Viewer
Comprehensive review of chronic stress pathways and the efficacy of behavioral stress reduction programs (bsrps) in managing diseases.

Share and Cite
Shchaslyvyi, A.Y.; Antonenko, S.V.; Telegeev, G.D. Comprehensive Review of Chronic Stress Pathways and the Efficacy of Behavioral Stress Reduction Programs (BSRPs) in Managing Diseases. Int. J. Environ. Res. Public Health 2024 , 21 , 1077. https://doi.org/10.3390/ijerph21081077
Shchaslyvyi AY, Antonenko SV, Telegeev GD. Comprehensive Review of Chronic Stress Pathways and the Efficacy of Behavioral Stress Reduction Programs (BSRPs) in Managing Diseases. International Journal of Environmental Research and Public Health . 2024; 21(8):1077. https://doi.org/10.3390/ijerph21081077
Shchaslyvyi, Aladdin Y., Svitlana V. Antonenko, and Gennadiy D. Telegeev. 2024. "Comprehensive Review of Chronic Stress Pathways and the Efficacy of Behavioral Stress Reduction Programs (BSRPs) in Managing Diseases" International Journal of Environmental Research and Public Health 21, no. 8: 1077. https://doi.org/10.3390/ijerph21081077
Article Metrics
Article access statistics, further information, mdpi initiatives, follow mdpi.
Subscribe to receive issue release notifications and newsletters from MDPI journals
Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Review Article
- Published: 11 May 2020
Using genetics for social science
- K. Paige Harden ORCID: orcid.org/0000-0002-1557-6737 1 &
- Philipp D. Koellinger ORCID: orcid.org/0000-0001-7413-0412 2
Nature Human Behaviour volume 4 , pages 567–576 ( 2020 ) Cite this article
7361 Accesses
76 Citations
503 Altmetric
Metrics details
- Politics and international relations
Social science genetics is concerned with understanding whether, how and why genetic differences between human beings are linked to differences in behaviours and socioeconomic outcomes. Our review discusses the goals, methods, challenges and implications of this research endeavour. We survey how the recent developments in genetics are beginning to provide social scientists with a powerful new toolbox they can use to better understand environmental effects, and we illustrate this with several substantive examples. Furthermore, we examine how medical research can benefit from genetic insights into social-scientific outcomes and vice versa. Finally, we discuss the ethical challenges of this work and clarify several common misunderstandings and misinterpretations of genetic research on individual differences.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
24,99 € / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
111,21 € per year
only 9,27 € per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout
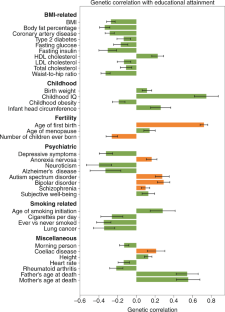
Similar content being viewed by others

Genetic determinism, essentialism and reductionism: semantic clarity for contested science

Moving back to the future of big data-driven research: reflecting on the social in genomics

Beyond the factor indeterminacy problem using genome-wide association data
Data availability.
The genetic correlations reported in Fig. 1 are based on publicly available GWAS summary statistics on LDHub ( http://ldsc.broadinstitute.org/ldhub/ ). The Health and Retirement Study data in Fig. 3 can be accessed via dbGaP and the University of Michigan.
Definition of social science . Merriam Webster Dictionary https://www.merriam-webster.com/dictionary/social%20science (Accessed 1 November 2018).
Turkheimer, E. Three laws of behavior genetics and what they mean. Curr. Dir. Psychol. Sci. 9 , 160–164 (2000).
Google Scholar
Polderman, T. J. C. et al. Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nat. Genet. 47 , 702–709 (2015).
CAS PubMed Google Scholar
Benjamin, D. J. et al. The promises and pitfalls of genoeconomics. Annu. Rev. Econ. 4 , 627–662 (2012).
Visscher, P. M. et al. 10 years of GWAS discovery: Biology, function, and translation. Am. J. Hum. Genet. 101 , 5–22 (2017).
CAS PubMed PubMed Central Google Scholar
Buniello, A. et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 47 D1, D1005–D1012 (2019).
Freese, J. The arrival of social science genomics. Contemp. Sociol. 47 , 524–536 (2018).
Comfort, N. Nature still battles nurture in the haunting world of social genomics. Nature 553 , 278–280 (2018).
Turkheimer, E. & Paige Harden, K. Behavior Genetic Research Methods. in Handbook of Research Methods in Social and Personality Psychology 159–187 (Cambridge University Press, 2014).
Kong, A. et al. The nature of nurture: Effects of parental genotypes. Science 359 , 424–428 (2018).
Koellinger, P. D. & Harden, K. P. Using nature to understand nurture: Genetic associations show how parenting matters for children’s education. Science 359 , 386–387 (2018).
Bycroft, C. et al. The UK Biobank resource with deep phenotyping and genomic data. Nature 562 , 203–209 (2018).
Servick, K. Can 23andMe have it all? Science 349 , 1472–1474, 1476–1477 (2015).
Duncan, L. E., Pollastri, A. R. & Smoller, J. W. Mind the gap: why many geneticists and psychological scientists have discrepant views about gene-environment interaction (G×E) research. Am. Psychol. 69 , 249–268 (2014).
PubMed PubMed Central Google Scholar
Reich, D. E. et al. Linkage disequilibrium in the human genome. Nature 411 , 199–204 (2001).
Abecasis, G. R. et al. The 1000 Genomes Project Consortium. An integrated map of genetic variation from 1,092 human genomes. Nature 491 , 56–65 (2012).
PubMed Google Scholar
Price, A. L. et al. Principal components analysis corrects for stratification in genome-wide association studies. Nat. Genet. 38 , 904–909 (2006).
Visscher, P. M., Brown, M. A., McCarthy, M. I. & Yang, J. Five years of GWAS discovery. Am. J. Hum. Genet. 90 , 7–24 (2012).
Chabris, C. F., Lee, J. J., Cesarini, D., Benjamin, D. J. & Laibson, D. I. The fourth law of behavior genetics. Curr. Dir. Psychol. Sci. 24 , 304–312 (2015).
Rapid GWAS of thousands of phenotypes for 337,000 samples in the UK Biobank. Neale Lab http://www.nealelab.is/blog/2017/7/19/rapid-gwas-of-thousands-of-phenotypes-for-337000-samples-in-the-uk-biobank (Accessed 6 November 2018).
Kyoko Watanabe, D.P. Atlas of GWAS Summary Statistics. GWAS Atlas ( 2017 ) . http://atlas.ctglab.nl/ (Accessed 6 November 2019).
Global Biobank Engine. https://biobankengine.stanford.edu/ (Accessed 6 November 2019).
Manolio, T. A. et al. Finding the missing heritability of complex diseases. Nature 461 , 747–753 (2009).
Sohail, M. et al. Polygenic adaptation on height is overestimated due to uncorrected stratification in genome-wide association studies. eLife 8 , e39702 (2019).
Berg, J. J. et al. Reduced signal for polygenic adaptation of height in UK Biobank. eLife 8 , e39725 (2019).
Haworth, S. et al. Apparent latent structure within the UK Biobank sample has implications for epidemiological analysis. Nat. Commun. 10 , 333 (2019).
Lawson, D. J. et al. Is population structure in the genetic biobank era irrelevant, a challenge, or an opportunity? Hum. Genet. 139 , 23–41 (2020).
Loh, P.-R. et al. Efficient Bayesian mixed-model analysis increases association power in large cohorts. Nat. Genet. 47 , 284–290 (2015).
Yang, J., Zaitlen, N. A., Goddard, M. E., Visscher, P. M. & Price, A. L. Advantages and pitfalls in the application of mixed-model association methods. Nat. Genet. 46 , 100–106 (2014).
Davies, N. M. et al. Within family Mendelian randomization studies. Hum. Mol. Genet. 28 R2 R170–R179, https://doi.org/10.1093/hmg/ddz204 (2019).
Bulik-Sullivan, B. et al. An atlas of genetic correlations across human diseases and traits. Nat. Genet. 47 , 1236–1241 (2015).
Lee, J. J., McGue, M., Iacono, W. G. & Chow, C. C. The accuracy of LD Score regression as an estimator of confounding and genetic correlations in genome-wide association studies. Genet. Epidemiol. 42 , 783–795 (2018).
Zheng, J. et al. LD Hub: a centralized database and web interface to perform LD score regression that maximizes the potential of summary level GWAS data for SNP heritability and genetic correlation analysis. Bioinformatics 33 , 272–279 (2017).
Lee, J. J. et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat. Genet. 50 , 1112–1121 (2018).
Gage, S. H., Davey Smith, G., Ware, J. J., Flint, J. & Munafò, M. R. G = E: what GWAS can tell us about the environment. PLoS Genet. 12 , e1005765 (2016).
Turley, P. et al. Multi-trait analysis of genome-wide association summary statistics using MTAG. Nat. Genet. 50 , 229–237 (2018).
Grotzinger, A. D. et al. Genomic structural equation modelling provides insights into the multivariate genetic architecture of complex traits. Nat. Hum. Behav. 3 , 513–525 (2019).
Baselmans, B. M. L. et al. Multivariate genome-wide analyses of the well-being spectrum. Nat. Genet. 51 , 445–451 (2019).
Loehlin, J. C. The Cholesky approach: A cautionary note. Behav. Genet. 26 , 65–69 (1996).
Torkamani, A., Wineinger, N. E. & Topol, E. J. The personal and clinical utility of polygenic risk scores. Nat. Rev. Genet. 19 , 581–590 (2018).
Dudbridge, F. Power and predictive accuracy of polygenic risk scores. PLoS Genet. 9 , e1003348 (2013).
Wray, N. R. et al. Research review: Polygenic methods and their application to psychiatric traits. J. Child Psychol. Psychiatry 55 , 1068–1087 (2014).
de Vlaming, R. et al. Meta-GWAS accuracy and power (MetaGAP) calculator shows that hiding heritability is partially due to imperfect genetic correlations across studies. PLoS Genet. 13 , e1006495 (2017).
Rietveld, C. A. et al. GWAS of 126,559 individuals identifies genetic variants associated with educational attainment. Science 340 , 1467–1471 (2013).
Okbay, A. et al. Genome-wide association study identifies 74 loci associated with educational attainment. Nature 533 , 539–542 (2016).
Wood, A. R. et al. Defining the role of common variation in the genomic and biological architecture of adult human height. Nat. Genet. 46 , 1173–1186 (2014).
Yengo, L. et al. Meta-analysis of genome-wide association studies for height and body mass index in ∼ 700000 individuals of European ancestry. Hum. Mol. Genet. 27 , 3641–3649 (2018).
Belsky, D. W. & Harden, K. P. Phenotypic annotation: Using polygenic scores to translate discoveries from genome-wide association studies from the top down. Curr. Dir. Psychol. Sci. 28 , 82–90 (2019).
Barcellos, S. H., Carvalho, L. S. & Turley, P. Education can reduce health differences related to genetic risk of obesity. Proc. Natl Acad. Sci. USA 115 , E9765–E9772 (2018).
Bansal, V. et al. Genome-wide association study results for educational attainment aid in identifying genetic heterogeneity of schizophrenia. Nat. Commun. 9 , 3078 (2018).
Belsky, D. W. et al. The genetics of success: how single-nucleotide polymorphisms associated with educational attainment relate to life-course development. Psychol. Sci. 27 , 957–972 (2016).
Young, A. I., Benonisdottir, S., Przeworski, M. & Kong, A. Deconstructing the sources of genotype-phenotype associations in humans. Science 365 , 1396–1400 (2019).
Mostafavi, H., Harpak, A., Conley, D., Pritchard, J. K. & Przeworski, M. Variable prediction accuracy of polygenic scores within an ancestry group. eLife 9 , e48376 (2020).
DiPrete, T. A., Burik, C. A. P. & Koellinger, P. D. Genetic instrumental variable regression: Explaining socioeconomic and health outcomes in nonexperimental data. Proc. Natl Acad. Sci. USA 115 , E4970–E4979 (2018).
Wertz, J. et al. Genetics of nurture: A test of the hypothesis that parents’ genetics predict their observed caregiving. Dev. Psychol. 55 , 1461–1472 (2019).
SSGAC Polygenic Score Data. Health and Retirement Study https://hrs.isr.umich.edu/news/ssgac-polygenic-score-data (Accessed 11 November 2019).
Young, A. I. et al. Relatedness disequilibrium regression estimates heritability without environmental bias. Nat. Genet. 50 , 1304–1310 (2018).
Davey Smith, G. & Hemani, G. Mendelian randomization: genetic anchors for causal inference in epidemiological studies. Hum. Mol. Genet. 23 R1 R89–R98 (2014).
Brumpton, B. et al. Within-family studies for Mendelian randomization: avoiding dynastic, assortative mating, and population stratification biases. bioRxiv https://doi.org/10.1101/602516 (2019).
O’Connor, L. J. & Price, A. L. Distinguishing genetic correlation from causation across 52 diseases and complex traits. Nat. Genet. 50 , 1728–1734 (2018).
Zhu, Z. et al. Causal associations between risk factors and common diseases inferred from GWAS summary data. Nat. Commun. 9 , 224 (2018).
Verbanck, M., Chen, C.-Y., Neale, B. & Do, R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 50 , 693–698 (2018).
Koellinger, P. D. & de Vlaming, R. Mendelian randomization: the challenge of unobserved environmental confounds. Int. J. Epidemiol. 48 , 665–671 (2019).
Black, S. E., Devereux, P. J. & Salvanes, K. G. Why the apple doesn’t fall far: Understanding intergenerational transmission of human capital. Am. Econ. Rev. 95 , 437–449 (2005).
Bates, T. C. et al. The nature of nurture: Using a virtual-parent design to test parenting effects on children’s educational attainment in genotyped families. Twin Res. Hum. Genet. 21 , 73–83 (2018).
Liu, H. Social and genetic pathways in multigenerational transmission of educational attainment. Am. Sociol. Rev. 83 , 278–304 (2018).
Belsky, D. W. et al. Genetic analysis of social-class mobility in five longitudinal studies. Proc. Natl Acad. Sci. USA 115 , E7275–E7284 (2018).
Barth, D., Papageorge, N. W. & Thom, K. Genetic endowments and wealth inequality. J. Polit. Econ. 128 , 1474–1522 (2020).
Holland, P. W. Statistics and causal inference. J. Am. Stat. Assoc. 81 , 945–960 (1986).
Rodgers, J. & Kohler, H.-P. The Biodemography of Human Reproduction and Fertility . (Springer, 2002).
Barban, N. et al. Genome-wide analysis identifies 12 loci influencing human reproductive behavior. Nat. Genet. 48 , 1462–1472 (2016).
Day, F. R. et al. Genomic analyses identify hundreds of variants associated with age at menarche and support a role for puberty timing in cancer risk. Nat. Genet. 49 , 834–841 (2017).
Day, F. R. et al. Physical and neurobehavioral determinants of reproductive onset and success. Nat. Genet. 48 , 617–623 (2016).
Mehta, D. et al. Evidence for genetic overlap between schizophrenia and age at first birth in women. JAMA Psychiatry 73 , 497–505 (2016).
Ni, G., Gratten, J., Wray, N. R. & Lee, S. H., Schizophrenia Working Group of the Psychiatric Genomics Consortium. Age at first birth in women is genetically associated with increased risk of schizophrenia. Sci. Rep. 8 , 10168 (2018).
Harden, K. P. et al. A behavior genetic investigation of adolescent motherhood and offspring mental health problems. J. Abnorm. Psychol. 116 , 667–683 (2007).
Beauchamp, J. P. Genetic evidence for natural selection in humans in the contemporary United States. Proc. Natl Acad. Sci. USA 113 , 7774–7779 (2016).
Kong, A. et al. Selection against variants in the genome associated with educational attainment. Proc. Natl Acad. Sci. USA 114 , E727–E732 (2017).
Rodgers, J. L. et al. Education and cognitive ability as direct, mediating, or spurious influences on female age at first birth: behavior genetic models fit to Danish twin data. Am. J.Sociol. 114 , S202–S232 (2008). Suppl.
Tropf, F. C. & Mandemakers, J. J. Is the association between education and fertility postponement causal? The role of family background factors. Demography 54 , 71–91 (2017).
Jocklin, V., McGue, M. & Lykken, D. T. Personality and divorce: a genetic analysis. J. Pers. Soc. Psychol. 71 , 288–299 (1996).
D’Onofrio, B. M., Eaves, L. J., Murrelle, L., Maes, H. H. & Spilka, B. Understanding biological and social influences on religious affiliation, attitudes, and behaviors: a behavior genetic perspective. J. Pers. 67 , 953–984 (1999).
Pilling, L. C. et al. Human longevity: 25 genetic loci associated in 389,166 UK biobank participants. Aging (Albany NY) 9 , 2504–2520 (2017).
CAS Google Scholar
Abdellaoui, A. et al. Genetic correlates of social stratification in Great Britain. Nat. Hum. Behav. 3 , 1332–1342 (2019).
Hill, W. D. et al. Molecular genetic contributions to social deprivation and household income in UK Biobank. Curr. Biol. 26 , 3083–3089 (2016).
Belsky, D. W. et al. Genetics and the geography of health, behaviour and attainment. Nat. Hum. Behav. 3 , 576–586 (2019).
van der Sluis, S., Posthuma, D. & Dolan, C. V. A note on false positives and power in G × E modelling of twin data. Behav. Genet. 42 , 170–186 (2012).
Duncan, L. E. & Keller, M. C. A critical review of the first 10 years of candidate gene-by-environment interaction research in psychiatry. Am. J. Psychiatry 168 , 1041–1049 (2011).
Ceci, S. J. & Papierno, P. B. The rhetoric and reality of gap closing: when the “have-nots” gain but the “haves” gain even more. Am. Psychol. 60 , 149–160 (2005).
Fletcher, J. M. Why have tobacco control policies stalled? Using genetic moderation to examine policy impacts. PLoS One 7 , e50576 (2012).
Goldberger, A. S. Heritability. Economica 46 , 327–347 (1979).
Jencks, C. Heredity, environment, and public policy reconsidered. Am. Sociol. Rev. 45 , 723–736 (1980).
Bronfenbrenner, U. & Ceci, S. J. Nature-nurture reconceptualized in developmental perspective: a bioecological model. Psychol. Rev. 101 , 568–586 (1994).
Dickens, W. T. & Flynn, J. R. Heritability estimates versus large environmental effects: the IQ paradox resolved. Psychol. Rev. 108 , 346–369 (2001).
Lykken, D. T., Bouchard, T. J. Jr., McGue, M. & Tellegen, A. Heritability of interests: a twin study. J. Appl. Psychol. 78 , 649–661 (1993).
Tucker-Drob, E. M. & Harden, K. P. Early childhood cognitive development and parental cognitive stimulation: evidence for reciprocal gene-environment transactions. Dev. Sci. 15 , 250–259 (2012).
Klahr, A. M., Thomas, K. M., Hopwood, C. J., Klump, K. L. & Burt, S. A. Evocative gene-environment correlation in the mother-child relationship: a twin study of interpersonal processes. Dev. Psychopathol. 25 , 105–118 (2013).
Scarr, S. & McCartney, K. How people make their own environments: a theory of genotype greater than environment effects. Child Dev. 54 , 424–435 (1983).
Wray, N. R. et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat. Genet. 50 , 668–681 (2018).
Wehby, G. L., Domingue, B. W. & Wolinsky, F. D. Genetic risks for chronic conditions: Implications for long-term wellbeing. J. Gerontol. A Biol. Sci. Med. Sci. 73 , 477–483 (2018).
Mallard, T. T., Harden, K. P. & Fromme, K. Genetic risk for schizophrenia is associated with substance use in emerging adulthood: an event-level polygenic prediction model. Psychol. Med. 49 , 2027–2035 (2019).
Okbay, A. et al. Genetic variants associated with subjective well-being, depressive symptoms, and neuroticism identified through genome-wide analyses. Nat. Genet. 48 , 624–633 (2016).
Wootton, R. E. et al. Evaluation of the causal effects between subjective wellbeing and cardiometabolic health: mendelian randomisation study. Br. Med. J. 362 , k3788 (2018).
Kevles, D.J. In the Name of Eugenics: Genetics and the Uses of Human Heredity . (Harvard University Press, 1995).
Rawls, J. A Theory of Justice . (Belknap Press, 1999).
Herrnstein, R.J. & Murray, C. The Bell Curve: Intelligence and Class Structure in American Life (Free Press, 1996).
Jensen, A. How much can we boost IQ and scholastic achievement. Harv. Educ. Rev. 39 , 1–123 (1969).
Yudell, M., Roberts, D., DeSalle, R. & Tishkoff, S. Taking race out of human genetics. Science 351 , 564–565 (2016).
Smedley, A. & Smedley, B. D. Race as biology is fiction, racism as a social problem is real: Anthropological and historical perspectives on the social construction of race. Am. Psychol. 60 , 16–26 (2005).
Henrich, J., Heine, S. J. & Norenzayan, A. The weirdest people in the world? Behav. Brain Sci. 33 , 61–83 (2010). discussion 83–135.
Martin, A. R. et al. Clinical use of current polygenic risk scores may exacerbate health disparities. Nat. Genet. 51 , 584–591 (2019).
Martin, A. R. et al. Human demographic history impacts genetic risk prediction across diverse populations. Am. J. Hum. Genet. 100 , 635–649 (2017).
Panofsky, A. & Donovan, J. Genetic ancestry testing among white nationalists: From identity repair to citizen science. Soc. Stud. Sci . https://doi.org/10.1177/0306312719861434 (2019).
Novembre, J. & Barton, N. H. Tread lightly interpreting polygenic tests of selection. Genetics 208 , 1351–1355 (2018).
Download references
Acknowledgements
We thank C. Burik for preparing Fig. 1 and the Social Science Genetic Association Consortium ( https://www.thessgac.org/ ) for Fig. 3 . P.D.K. was financially supported by an ERC consolidator grant (647648 EdGe). K.P.H. was supported by the Jacobs Foundation, the Templeton Foundation and Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD) grants R01-HD083613 and 5-R24-HD042849 (to the Population Research Center at the University of Texas at Austin). The funders had no role in the conceptualization, preparation or decision to publish this work.
Author information
Authors and affiliations.
Department of Psychology, University of Texas at Austin, Austin, Texas, USA
K. Paige Harden
Department of Economics, Vrije Universiteit Amsterdam, Amsterdam, The Netherlands
Philipp D. Koellinger
You can also search for this author in PubMed Google Scholar
Corresponding authors
Correspondence to K. Paige Harden or Philipp D. Koellinger .
Ethics declarations
Competing interests.
The authors declare no competing interests.
Additional information
Editor recognition statement: Primary handling editor: Stavroula Kousta.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Reprints and permissions
About this article
Cite this article.
Harden, K.P., Koellinger, P.D. Using genetics for social science. Nat Hum Behav 4 , 567–576 (2020). https://doi.org/10.1038/s41562-020-0862-5
Download citation
Received : 31 January 2019
Accepted : 16 March 2020
Published : 11 May 2020
Issue Date : 01 June 2020
DOI : https://doi.org/10.1038/s41562-020-0862-5
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
This article is cited by
Could interventions on physical activity mitigate genomic liability for obesity applying the health disparity framework in genetically informed studies.
- Moritz Herle
- Andrew Pickles
- Bianca L. De Stavola
European Journal of Epidemiology (2023)
The impact of digital media on children’s intelligence while controlling for genetic differences in cognition and socioeconomic background
- Bruno Sauce
- Magnus Liebherr
- Torkel Klingberg
Scientific Reports (2022)
Delayed tracking and inequality of opportunity: Gene-environment interactions in educational attainment
- Antonie Knigge
- Dorret I. Boomsma
npj Science of Learning (2022)
Genetics of cognitive performance, education and learning: from research to policy?
- Peter M. Visscher
Paternal Incarceration, Polygenic Scores, and Children’s Educational Attainment
- Breanna Clark
Journal of Developmental and Life-Course Criminology (2022)
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.

Research shows our bodies go through rapid changes in our 40s and our 60s
For many people, reaching their mid-40s may bring unpleasant signs the body isn’t working as well as it once did. Injuries seem to happen more frequently. Muscles may feel weaker.
A new study, published Wednesday in Nature Aging , shows what may be causing the physical decline. Researchers have found that molecules and microorganisms both inside and outside our bodies are going through dramatic changes, first at about age 44 and then again when we hit 60. Those alterations may be causing significant differences in cardiovascular health and immune function.
The findings come from Stanford scientists who analyzed blood and other biological samples of 108 volunteers ages 25 to 75, who continued to donate samples for several years.
“While it’s obvious that you’re aging throughout your entire life, there are two big periods where things really shift,” said the study’s senior author, Michael Snyder, a professor of genetics and director of the Center for Genomics and Personalized Medicine at Stanford Medicine. For example, “there’s a big shift in the metabolism of lipids when people are in their 40s and in the metabolism of carbohydrates when people are in their 60s.”
Lipids are fatty substances, including LDL, HDL and triglycerides, that perform a host of functions in the body, but they can be harmful if they build up in the blood.
The scientists tracked many kinds of molecules in the samples, including RNA and proteins, as well as the participants’ microbiomes.
The metabolic changes the researchers discovered indicate not that people in their 40s are burning calories more slowly but rather that the body is breaking food down differently. The scientists aren’t sure exactly what impact those changes have on health.
Previous research showed that resting energy use, or metabolic rate , didn’t change from ages 20 to 60. The new study’s findings don't contradict that.
The changes in metabolism affect how the body reacts to alcohol or caffeine, although the health consequences aren’t yet clear. In the case of caffeine, it may result in higher sensitivity.
It’s also not known yet whether the shifts could be linked to lifestyle or behavioral factors. For example, the changes in alcohol metabolism might be because people are drinking more in their mid-40s, Snyder said.
For now, Snyder suggests people in their 40s keep a close eye on their lipids, especially LDL cholesterol.
“If they start going up, people might want to think about taking statins if that’s what their doctor recommends,” he said. Moreover, “knowing there’s a shift in the molecules that affect muscles and skin, you might want to warm up more before exercising so you don’t hurt yourself.”
Until we know better what those changes mean, the best way to deal with them would be to eat healthy foods and to exercise regularly, Snyder said.
Dr. Josef Coresh, founding director of the Optimal Aging Institute at the NYU Grossman School of Medicine, compared the new findings to the invention of the microscope.
“The beauty of this type of paper is the level of detail we can see in molecular changes,” said Coresh, a professor of medicine at the school. “But it will take time to sort out what individual changes mean and how we can tailor medications to those changes. We do know that the origins of many diseases happen in midlife when people are in their 40s, though the disease may occur decades later.”
The new study “is an important step forward,” said Dr. Lori Zeltser, a professor of pathology and cell biology at the Columbia University Vagelos College of Physicians and Surgeons. While we don’t know what the consequences of those metabolic changes are yet, “right now, we have to acknowledge that we metabolize food differently in our 40s, and that is something really new.”
The shifts the researchers found might help explain numerous age-related health changes, such as muscle loss, because “your body is breaking down food differently,” Zeltser said.
Linda Carroll is a regular health contributor to NBC News. She is coauthor of "The Concussion Crisis: Anatomy of a Silent Epidemic" and "Out of the Clouds: The Unlikely Horseman and the Unwanted Colt Who Conquered the Sport of Kings."
An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- HHS Author Manuscripts
Overview of Behavioral Genetics Research for Family Researchers
Diana samek.
Department of Psychology, N218 Elliot Hall, 75 East River Road, Minneapolis, MN 55108 ( [email protected] ).
Martha Rueter
* Department of Family Social Science, 290 McNeal Hall, University of Minnesota, 1985 Buford Avenue, St. Paul, MN 55108 ( ude.nmu@reteurm ).
Bibiana Koh
** Department of Social Work, Augsburg College, 2211 Riverside Avenue, Minneapolis, MN 55454 ( ude.grubsgua@hok ).
This article provides an overview of the methods, assumptions, and key findings of behavioral genetics methodology for family researchers with a limited background. We discuss how family researchers can utilize and contribute to the behavioral genetics field, particularly in terms of conducting research that seeks to explain shared environmental effects. This can be done, in part, by theoretically controlling for genetic confounds in research that seeks to determine cause-and-effect relationships among family variables and individual outcomes. Gene–environment correlation and interaction are especially promising areas for the family researcher to address. Given the methodological advancements in the field, we also briefly comment on new methods in molecular genetics for studying psychological mental health disorders.
The debate between proponents of nature ( Scarr & McCartney, 1983 ) versus nurture ( Collins, Maccoby, Steinberg, Hetherington, & Bornstein, 2000 ) as an explanation for social behavior is long standing, well documented, and continues today (see Dar-Nimrod & Heine, 2011 ; Rende & Slomkowski, 2009 ; Turkheimer, 2011 ). In family social science, we typically focus on nurture effects and can sometimes overlook possible biological and/or genetic factors when examining family processes ( D’Onofrio & Lahey, 2010 ). This notion has been challenged by the broad conclusions that have come out of the behavioral genetics field claiming that additive genetic effects, rather than shared environmental effects, explain the most variance in traits and outcomes that we typically study (e.g., Plomin & Daniels, 1987 ). Those new to behavioral genetics research may not understand what the definitions of shared environmental effects are, how assumptions underlying behavioral genetics methods affect the interpretation of those findings, or how behavioral genetics findings challenge many of the original broad reports of little family influence on behavior ( Burt, 2009 ; Horwitz & Neiderhiser, 2011 ; Legrand, Keyes, McGue, Iacono, & Krueger, 2008 ).
Consistent with a recent call for family social science to control for genes and other biological influences ( D’Onofrio & Lahey, 2010 ; Horwitz & Neiderhiser, 2011 ), we believe that behavioral genetics findings have profound implications for family researchers. We address these implications by reviewing the methods, assumptions, and findings from the behavioral genetics field. This article is written by family researchers for family researchers who are not familiar with this line of work.
The majority of this article focuses on quantitative behavioral genetics research in terms of theory, methods, and key findings. We focus on future research questions that we believe are particularly appropriate for family researchers to address, including the identification of specific shared environmental variants in the development of adolescent substance use and related adjustment outcomes. We cover research on main effects, gene–environment correlation ( r GE), and gene–environment interaction (G×E). The work of behavioral genetics has led to research in molecular genetics that aims to identify specific genetic variants (main and G×E effects) in relation to psychopathology. Because of this advancement, we believe it is necessary to also briefly review recent molecular genetic methods and then comment on the relevance of this research to family scholars. Finally, we should mention that this article is not intended to provide a comprehensive review of the field; rather, it is a theoretical review of areas in behavior genetics that seem especially promising for family researchers to address. For a more comprehensive review, readers should refer to Bazzett (2008) ; Dick, Latendresse, and Riley (2011) ; Kim (2009) ; and/or Plomin, DeFries, McClearn, and McGuffin (2008) , which helped guide much of this article.
What is Behavioral Genetics?
Behavioral genetics is a branch of psychology that attempts to allocate and explain genetic and environmental contributions to human (and animal) behavior (Behavior Genetics Association, n.d.). Pursuing such knowledge involves defining behavioral outcomes (called phenotypes) and measuring the corresponding genetic influences (called genotypes).
Quantitative behavioral genetics methodology theoretically and statistically controls for genetic effects. “Theoretical control” refers to simply comparing study results (e.g., the association between parenting and child externalizing behaviors) across different, genetically informed family subsamples (e.g., monozygotic versus dizygotic twins, full biological siblings versus adopted or step siblings). Statistically, this methodology controls for genetic effects through a simple variance–covariance decomposition (called biometric modeling). In the next section, we more fully describe biometric modeling and the assumptions used to test behavioral genetic models.
A Look at the Methods in Quantitative Behavioral Genetics
Typically, one sees something like Figure 1 in quantitative behavioral genetics papers. Behavioral geneticists would describe this figure as a phenotype being “decomposed” into three basic categories: additive genetic (A), shared environmental (C), and nonshared environmental (E) influences. This means that some behavioral outcome of interest (i.e., the phenotype) will be partialed out in a regression-type framework to describe basic genetic and environmental effects in aggregate. To understand the basics of how these effects are calculated, it helps to first describe the sampling strategy used in such designs, as well as to give some formal definitions.

Variance components are represented by capital letters (additive genetic: A; shared environmental: C; nonshared environmental effects: E) using a twin design. Path coefficients are represented by lower case letters ( a, c, e ). Squaring the path coefficients (e.g., a 2 ) represents the percentage of variance in the trait by the variance component.
Sampling Strategy
A quantitative behavioral genetic study typically uses a genetically informed sibling-pair design ( Plomin et al., 2008 ). This type of study recruits a sample of families with at least two siblings who share varying proportions of genes (either within or across families). A twin study is most often used. In the twin design, researchers compare results across monozygotic twins (who share 100% of their DNA) and dizygotic twins (who share 50% of their DNA, on average; e.g., Iacono & McGue, 2002 ). Another type of behavioral genetics study design is an adoption study (e.g., McGue et al., 2007 ). In this type of study, researchers compare siblings who are the biological offspring of the same parents (i.e., they are full siblings and share 50% of their DNA) to adopted siblings who are not biologically related to their parents or to one another (i.e., they share 0% of their DNA). In a full adoption study, the biological parents of the adopted offspring are also included ( Leve, Neiderhiser, Scaramella, & Reiss, 2008 ). Correlation comparisons can be made across stepsiblings (who share 0% of their DNA), or half siblings (who share 25% of their DNA). Other designs are used, such as the Children of Twins (CoT) design ( D’Onofrio, 2005 ); however, they are somewhat less common.
Formal Definitions
Additive genetic influences ( a 2 , often reported as h 2 ) provide a ballpark estimate of a trait’s heritability ( Visscher, Hill, & Wray, 2008 ) and refer to the causes of two sibling’s similarity as a result of additive genetic influence. The shared (or common rearing) environmental influence ( c 2 ) accounts for similarities between siblings that are not due to genetics ( Burt, 2009 ). Typically, this refers to the family environment, but it is not necessary or sufficient for family variance to only be determined by shared environmental influence. This also includes any other environments siblings share, such as school, peers, and neighborhoods. Finally, nonshared environmental influences ( e 2 ) are defined as unique environments, and as situational or contextual factors that make siblings different. Theoretically, this refers to anything in the environment that makes siblings different, including parental differential treatment or unique peer groups. Empirically, this also includes any measurement error.
Behavioral Genetics Statistical Strategy: Biometric Modeling
The variance components defined earlier (A, C, E) and path coefficients ( a, c, e ) are statistically estimated using biometric modeling techniques (Neale, 2008; Plomin et al., 2008 ). Squaring the standardized path coefficients gives the percentage of variance explained on the specific trait examined (i.e., a 2 = heritability, c 2 = shared environment contribution, e 2 = nonshared environmental contribution). Figure 1 shows an example of a biometric model. As Figure 1 describes, biometric modeling is based on structural equation modeling, which utilizes the concept of latent factors. Latent factors are variables that are “hidden,” or difficult to measure. In biometric modeling, the latent factors refer to genetic and environmental contributions to a phenotype of interest (A, C, and E, which are assumed to be 1.0).
Figure 1 shows two squares that depict an observed phenotype, one for “Sibling 1” and the other for “Sibling 2.” The circles refer to the latent factors (which are the variance components representing effects of additive genetic, A ; shared environmental, C ; and nonshared environmental, E ). The arrows pointing from the circles to the phenotypes represent the estimation of the extent to which the variance component explains variance in the phenotype. The double-headed arrows connecting the two A variance components across siblings denotes a correlation ( r ), which is set a priori within sibling subsamples depending on their proportion of shared genes. This model is illustrated for a study of a single trait (phenotype) observed in monozygotic twins ( r MZ ) and dizygotic twins ( r DZ ). The A-to-A correlation for monozygotic twins is fixed to 1.0 (because they share 100% of their genes), and for dizygotic twins, it is fixed to .50 (because they share 50% of their genes). The correlation between C and C denotes the proportion of the shared environment the siblings have in common; it is set to 1.0 in both subsamples because biometric modeling assumes that monozygotic and dizygotic twins have the same environment (we will return to this assumption in a moment).
Finally, there is no a priori expectation for the magnitude of the correlation between the nonshared environment components (i.e., E). Statistically, E is anything left over after A and C have been partialed out, including measurement error; in fact, E is akin to the residual (or error term) in a simple regression equation. Theoretically, and consistent with our earlier definition, E refers to anything in the environment that makes siblings different (including unique environments and/or contextual and situational factors).
Given the gap between what is statistically measured in a biometric model and what is theoretically implied or inferred, care should be taken when interpreting E. In fact, studies have produced little consistent evidence of what is specifically accounted for by nonshared environmental influence (for a meta-analysis, see Turkheimer & Waldron, 2000 ). Not being able to identify specific nonshared environmental variants has led some to believe that the nonshared environment largely acts through idiosyncrasies within individuals and families that are difficult to measure ( Turkheimer & Waldron, 2000 ). However, because identical twins share 100% of the same genes and rearing environment, an analysis of discordant identical twins may be the most direct test of nonshared environmental influence ( Plomin, 2011 ). For example, Burt, McGue, Iacono, and Krueger (2006) analyzed monozygotic twins discordant on exposure to an event (e.g., parental treatment) in relation to later outcomes (e.g., externalizing). Burt et al. found that differential parent–child conflict predicted differential externalizing 3 years later, but only in the twins who were the most discordant in parent–child conflict. This match in discordance (if one twin had greater parent–child conflict than the other twin, then that twin was more likely to have greater externalizing problems than the other twin) suggests nonshared environmental mediation for the most extreme difference in parental differential treatment. If there was a mismatch in concordance (if one twin had greater parent–child conflict but both twins had a similar level of externalizing problems), this would suggest common genetic factors explain this relationship.
Biometric Modeling: The Assumptions
The interpretation of most statistical analyses relies on several assumptions. Assumptions can sometimes be easily glossed over or ignored. However, accurate interpretation of statistical findings is best done within an understanding of the analyses’ assumptions, and interpretation of biometric modeling results is no exception. In this section, we review three major biometric modeling assumptions.
The first is the assumption of equality in environments between subsample comparisons. This is a critical assumption in both twin and adoption designs that has sometimes been challenged. Regarding twin designs, monozygotic twins report being treated more similarly than do dizygotic twins during childhood ( Borkenau, Riemann, Angleitner, & Spinath, 2002 ). Monozygotic twins also tend to spend more time together and have more similar peer groups compared to dizygotic twins ( Kendler & Gardner, 1998 ). In contrast, monozygotic-twin environmental similarity may be due to genetic rather than environmental influences ( Loehlin & Nichols, 1976 ). In other words, monozygotic twins may spend more time together and share more friends than dizygotic twins as a result of their genetic similarity rather than a result of having a different environment.
The equality assumption is also challenged in adoption designs. For example, adoptive parents tend to have higher socioeconomic status (SES), marital stability, and better mental health than parents with biological offspring, which are used as comparisons ( McGue et al., 2007 ). Given this restriction of range, shared environmental effects can be deflated ( Stoolmiller, 1999 ). Yet McGue et al. (2007) report that the range restriction in externalizing problems and SES had little effect on adoptive-sibling correlations for drug use, IQ, or delinquency.
The second major behavioral genetics assumption is that biometric modeling accounts only for additive genetic effects ( Plomin, Haworth, & Davis, 2009 ). In general, the additive genetic effect refers to the effect of what varies in our genome, added up over multiple locations across the genome. However, not all genetic influence is additive (e.g., dominance, epistasis). Whereas additive genetic influence refers to the affect of alleles, one by one, from all over the genome, dominance refers to the effect of one allele depending on the other in the same location in the genome. For example, phenylketonuria (PKU) operates with a dominant-recessive influence because affected offspring must have two copies of the PKU allele (if they have only one copy, they can be carriers and pass down that influence to their own children). Much like dominance, epitasis refers to the affect of one allele on another but at multiple locations across the genome. It has been argued that most psychological traits appear to have an additive, polygenic effect ( Plomin et al., 2009 ). It is important to note though, that when we speak about heritability within an additive genetic framework, we are referring only to narrow, and not broad-sense heritability (broad-sense accounts for all genotypic variance; Hamilton, 2009 ).
Main-effects biometric modeling also assumes that there is no correlation or interaction between genetic and environmental effects. In fact, there has been an increasing amount of work done on both of these concepts, referred to gene–environment interplay ( Horwitz & Neiderhiser, 2011 ; Moffitt, Caspi, & Rutter, 2005 ; Neiderhiser, 2011 ). In their landmark article, Scarr and McCartney (1983) describe three basic types of gene–environment correlations ( r GE): (1) evocative r GE (e.g., parents respond to children in the same family differently as a result of their different genotypes); (2) active r GE (e.g., children actively seek out environments that fit their phenotypes), and (3) passive r GE (e.g., parents’ genotypes influence both their own and their child’s behaviors). These explanations have been utilized when trying to make sense of why some traits are heritable, such as parent–child relationship quality (e.g., McGue, Elkins, Walden, & Iacono, 2005 ). Moreover, there are statistical tests for gene–environment interaction ( Purcell, 2002 ), and research in this area is extant (we cover some key points in this area of research later, after we finish covering assumptions and main effects findings).
Finally, biometric modeling assumes that dizygotic twins (or full siblings) share exactly 50% of their DNA, but this is actually an average. Full siblings can share about 40–60% of their DNA ( Visscher et al., 2006 ), and this variation can affect the accuracy of biometric estimates. Also, recall that human DNA is 99.5% identical across individuals. Thus, only a small proportion of DNA varies across family members, and it is those small variations that are being estimated when accounting for genetic differences across siblings.
In general, the assumptions used in behavioral genetics designs do not appear to be grossly violated; however, it is important to remember these assumptions in reading any behavioral genetics article. The best way to understand whether a violation of assumptions greatly influences study findings is (1) to find out if results have been replicated across a variety of behavioral genetics designs (e.g., twin and adoption studies) and (2) to be mindful of the concept of gene–environment interplay. Interpretations of the heritability of traits can be interpreted in a gene–environment correlation framework; if gene–environment interactions are not specifically tested, they may be relevant in the development of that trait as well. Also, (3) it is useful to find out whether the equal-environments assumption has been examined in each sample to see how much it matters to the phenotypes of interest (e.g., Kendler & Gardner, 1998 ; McGue et al., 2007 ). Finally, (4) recall that while most psychological traits are additive in nature, other genetic effects may be relevant as well.
Quantitative Behavioral Genetics: Major Findings
Now that we have laid the basic groundwork of behavioral genetics methods, definitions, and assumptions, we review key behavioral genetics research findings and explore their implications for family researchers. The goal of this discussion is to further evaluate the sources of a 2 , c 2 , and e 2 on the particular traits (observed phenotypes, such as personality and antisocial behavior) based on the current literature. Our primary focus is on the shared environment ( c 2 ), but we also cover additive genetic ( a 2 ) and nonshared environmental (e 2 ) effects. We focus on the shared environment because, given that we are considered “experts” on the family, we believe that family researchers could greatly contribute to this topic by conducting research that explains specific shared environmental variants.
Traits with negligible shared environmental effects
Many behavioral genetics studies have yielded additive genetic effects in a variety of traits, including aggression ( DiLalla, 2002 ), personality ( Bouchard & McGue, 2003 ), eating disorders ( Bulik, Sullivan, Wade, & Kendler, 2000 ), feelings and attitudes ( McGue, Sharma, & Benson, 1996 ), political attitudes and behaviors ( Hatemi, Dawes, Frost-Keller, Settle, & Verhulst, 2011 ), cognitive abilities and psychopathology ( Plomin & Rende, 1990 ), and even family relationship quality ( Elkins, McGue, & Iacono, 1997 ; McGue et al., 2005 ). Here we focus on two phenotypes whose small shared environmental effects have been broadly and exhaustively replicated: personality and mental health disorders.
On the basis of extant behavioral genetics personality research, the shared environment appears to account for almost no variance in personality. Additive genetic effects explain between 39% and 58%, and nonshared environmental effects account for 40% to 56% of variance in personality ( Bouchard & McGue, 2003 ; Tellegen et al., 1988 ). Confidence in these estimates is established through their replication across samples (e.g., McGue, Bouchard, Iacono, & Lykken, 1993 ; Plomin, Owen, & McGuffin, 1994 ) using various personality structures including a three-factor model ( Tellegen et al., 1988 ), a five-factor model ( Jang, Livesley, & Vernon, 1996 ; Loehlin, McCrae, Costa, & John, 1998 ; Yamagata et al., 2006 ), and a general factor of personality ( Rushton, Bons, & Hur, 2008 ). Overall, the existing literature suggests that genetic effects can explain a substantial portion of personality traits. The presence of little to no shared environment effects and substantial nonshared effects, which include measurement error, suggests that either the shared environment has little effect on personality development or that relevant shared environment components of personality are not identified due to things like gene–environment correlation or interaction.
Similar patterns of genetic and environmental estimates have been found for mental disorders (for a review, see Kendler & Prescott, 2006 ). For example, using a pooled sample of national registries, Lichtenstein et al. (2009) found that less than 5% of variance in schizophrenia was attributed to the shared environment, and nearly 64% was attributed to additive genetic effects. A similar pattern was found for bipolar disorder (59% additive genetic, 3% shared environment). In fact, the comorbidity between these disorders is predominately explained by additive genetic effects, which suggests a common genetic cause for both disorders ( Lichtenstein et al., 2009 ).
Taken together, these findings imply that if we are interested in the etiology of personality and serious psychiatric disorders, then examining shared environmental effects in isolation may not be a promising approach. An alternative approach may be to examine family effects that contribute to nonshared environmental variance, because nonshared effects on personality and psychiatric disorders appear to be as substantial as additive genetic effects. However, this notion has led to few significant findings. For example, Pike, McGuire, Hetherington, Reiss, and Plomin (1996) aimed to detect nonshared environment covariance in the relationship between parenting and adolescent antisocial behavior and depression, but they found negligible effects for the environment and greater effects for genetic mediation in this relationship. In their well-known meta-analysis, Turkheimer and Waldron (2000) found little evidence for replicated nonshared environmental findings. More recent approaches to studying the environment, including attention to appropriate measurement tools, the use of advanced statistical procedures, and consideration of gene–environment interplay seem especially promising ( Horwitz & Neiderhiser, 2011 ; Loehlin, 2010 ; Turkheimer & Waldron, 2000 ). We return to the concept and methods after our discussion of main effects.
Traits with moderate to substantial shared environmental effects
In addition to traits with large additive genetic effects (e.g., personality), there are also traits with moderate to substantial shared environmental effects. One of the first articles to show moderate shared environmental effects on a variety of traits was by McGue et al. (1996) . Significant correlations between adoptive siblings were found for the behavioral traits (e.g., externalizing, antisocial behavior, prosocial behavior), but consistent with Tellegen et al.’s (1988) findings, not for personality traits and affective traits (e.g., internalizing, negative emotionality). Remember, according to behavioral genetics assumptions, because adopted siblings are not genetically related (they are selected as such for a behavioral genetics design), any correlation between them refers to similarity that is attributed to the shared environment. The shared environment explained roughly 10% of the variance in the behavioral traits, which implies a modest shared environmental effect. These findings were consistently found across several studies. A recent meta-analysis of shared environmental effects ( Burt, 2009 ) found that shared environment effects ranged from 10% to 16% on behavioral disinhibition traits (e.g., conduct disorder, oppositional defiant disorder, anxiety, depression). Additive genetic and nonshared environmental contributions to these childhood disorders were still substantial, ranging from 26% to 59%. In contrast, shared environmental effects increased to nearly 30% when considering important covariates such as age and gender.
There are also moderate shared environmental effects on IQ. For example, Buchanan, McGue, Keyes, and Iacono (2009) reported estimates of 19% for shared environmental, 65% for additive genetic, and 16% for nonshared environmental influences. Demonstrating a potential specific influence of SES on this shared environmental effect, Duyme, Dumaret, and Tomkiewicz (1999) reviewed 65 files of deprived adopted children (who are not genetically related to their adoptive parents) who were given IQ tests pre- and post-placement. These adopted children were determined to be deprived because of their experience of abuse or neglect, were adopted at older ages, and had IQs of less than 86. They documented an increase in IQ of more than 10 points after children were placed with their adoptive families. This suggests that merely being placed with their new adoptive families helped explain the increase in IQ points (which suggests shared environmental influences). Moreover, IQ increased more in higher-SES families, which suggests a specific shared environmental influence of SES on IQ in these adoptive families (because SES is a family-level variable). On average, IQ increased 7 points in low-SES adoptive families, 15 points in middle-SES adoptive families, and nearly 20 points in high-SES adoptive families. These results imply that SES may at least partially explain the shared environmental effect on IQ.
Shared environment effects are in fact quite substantial for behaviors such as adolescent substance use, generally explaining 30–60% of variance in adolescent substance use ( McGue, Elkins, & Iocono, 2000 ). In McGue et al.’s (2000) study, the additive genetic contribution to substance use in adolescence was approximately 13%, and the nonshared environmental contribution was approximately 26%, thus indicating important contributions for both. Substance-use results of this magnitude have been replicated (see Hopfer, Crowley, & Hewitt, 2003 ; Lynskey, Agrawal, & Health, 2010 ; Rende & Slomkowski, 2009 ) and call attention to the importance of furthering our understanding of shared environmental effects on adolescent substance use specifically.
The measurement of phenotypes is essential to consider. For example, we know that observer-rated behavior may be a better measure of shared environmental effects than self-reported parent–child relationship quality. Using a twin sample of toddlers, Deater-Deckard (2000) found that shared environment effects were better explained by observer ratings of children’s difficult behavior than by parent ratings. In this study, parents’ ratings of positive and negative child behavior showed genetic and nonshared environment mediation in child externalizing outcomes. In contrast, observer ratings showed shared environment mediation to those outcomes. Burt, Klahr, Rueter, McGue, and Iacono (2011) also found that observer ratings accounted for more shared environmental variance (31%) than other informant ratings, including maternal report (23%) and self-report (20%).
More Complex Analyses in Quantitative Behavioral Genetics
So far, we have determined that shared environmental effects are important, particularly for children and adolescents, and especially for outcomes and attributes such as IQ, externalizing behaviors, and substance use. Discovering the specific attributes of the shared environment that influence phenotypes is a necessary next step to further our knowledge of the etiology of these phenotypes. In other words, these findings point to something in the shared environment that may affect children who act out or explain why adolescents use substances. If we can identify the antecedent(s), these findings could help inform prevention and intervention research.
To move beyond this basic descriptive research, it is necessary to use more complex methods that examine gene–environment interplay ( Horwitz & Neiderhiser, 2011 ; Moffitt et al., 2005 ; Neiderhiser, 2011 ). There are at least two approaches to doing this. The first decomposes the correlations between family processes and child outcomes ( r GE), and the second models gene–environment interactions (G×E). Here we review and comment on literature that has examined both approaches.
Behavioral genetics: Decomposing correlations between family processes and child outcomes
Much like the methods used in a basic biometric model of a phenotype (also known as a univariate decomposition), a bivariate or multivariate biometric decomposition can also be used via behavioral genetic designs. In this type of model, in addition to the variance of each phenotype getting decomposed into genetic and environmental contributions (described in Figure 1 ), the covariance between phenotypes is also decomposed into genetic and environmental contributions. Thus, these models can estimate environmental versus genetic mediation in the association between two phenotypes.
Previous research using these multivariate biometric models has found that the relationship between parenting and child externalizing behavior is at least partly environmental in nature ( Burt, McGue, Krueger, & Iacono, 2007 ; Klahr, McGue, Iacono, & Burt, 2011 ; Neiderhiser, Pike, Hetherington, & Reiss, 1998 ). For example, Burt et al. (2007) determined that roughly 19% of the total variance in delinquency was explained by the shared environment (a fairly substantial amount of variance). Burt et al. additionally tested whether parent–child relationship quality (e.g., conflict, involvement) influenced child delinquency through the shared environment using multivariate decomposition. Indeed, they found that 2.8% of the variance in delinquency was accounted for by parenting variables and that those parenting variables explained 15% of the total shared environmental effect on delinquency.
Later research has shown additional shared environmental influences on associations between the family environment and child outcomes. For example, Shelton et al. (2008) also found shared environmental influences on the association between maternal warmth and adolescent conduct problems, further replicating Burt et al.’s (2007) report. Yet the proportion of shared environmental influence explained is still rather low in these reports, which suggests that further research is needed.
It has also been argued that family environment variables are too broadly defined (i.e., the measures need more theoretical guidance), which may be one reason we see less of an environmental influence on these family environment variables in behavioral genetics research. To challenge this, Latendresse, Rose, Viken, Pulkkinen, Kaprio, and Dick (2010) analyzed several measures of parenting and found some measures had greater shared environmental effects (parental discipline, relational tension) than others (autonomy granting, parental knowledge). Moreover, they found that parental knowledge and warmth alone explained 6–32% of the total shared environmental effects on adolescent drinking. These findings suggest that parent–child relationship quality may be an important contributor to shared environmental effects. However, more research is needed to understand what other aspects of the environment may contribute to shared environmental effects.
Family researchers can contribute to this unfolding of shared environmental effects by asking questions similar to the following: What are the specific characteristics of the shared environment that contribute to traits and outcomes? What is a theory of the shared environment? What is it about the shared environment that matters and why?
For example, following a growing body of research, it appears (and not surprisingly so) that much more than parenting influences child and adolescent outcomes within the family framework. Siblings, in particular, appear to be key influences on adolescents. In fact, while some research has found shared environmental influences of parenting (e.g., Burt et al., 2007 ; Latendresse et al., 2010 ), the magnitude of shared environmental effects appears to be greater for characteristics of sibling relationship quality than for parent–child relationship quality ( Neiderhiser, Reiss, & Hetherington, 2007 ). Conversely, parent–child relationship quality influences children more (than sibling relationship quality) through genetic influences ( Bussell et al., 1999 ; Fagan & Najman, 2005 ; Feinberg, Neiderhiser, Howe, & Hetherington, 2001 ; McGue & Iacono, 2009 ; Pike et al., 1996 ).
This “genetic mediation” of parenting on children’s outcomes is usually interpreted through a gene–environment correlation framework ( Burt, 2011 ; Horwitz & Neiderhiser, 2011 ; Scarr & McCartney, 1983 ). Specifically, an adolescent’s genotype may, in some way, be associated with parental behavior. This is consistent with the bidirectional parent–child relationship often employed in family theories, such that parents respond to their children on the basis of their unique, genetically influenced personality and behavior. This example is defined as evocative r GE. Another type of r GE is active; in this, adolescents are seeking out their own environments to match their genetic predispositions (e.g., antisocial adolescents wish to be less involved with their parents and more involved with antisocial peers). A final type of r GE is passive; in this, parents pass down their own genetic predispositions while simultaneously providing environmental influence (e.g., parents with histories of antisocial behavior transmit genes to their children and are less involved with their children).
Given how close siblings are in age, it has been argued that siblings may be important socializing influences on outcomes related specifically to substance use ( Lynskey et al., 2010 ). For example, using a genetically informed sample (full biological siblings and adopted siblings), Samek and Rueter (2011) found that sibling similarity in substance use was not significantly different across genetically related versus unrelated pairs and that feeling close to an elder sibling reduced the younger sibling’s overall substance-use behavior 3.5 years later. This, along with other research on sibling relationship quality using quantitative behavioral genetics designs ( Slomkowski, Rende, Novak, Lloyd-Richardson, & Niaura, 2005 ), suggests that the shared environment may largely pertain to the sibling relationship context.
Behavioral genetics: Gene–environment interactions
In an interaction, the strength of the correlation between the independent variable (X) and the dependent variable (Y) changes when considering the influence of a third variable, a moderator. For example, if the association between X and Y is stronger among fathers than among mothers, parent gender would be said to moderate the association between X and Y. Gene–environment interactions (G×E) are defined as genetic sensitivity to the environment ( Purcell, 2002 ); they help explain why people who share the same environmental experience have different consequences to events. G×E modeling detects whether the contributions of genetic, shared, and nonshared environmental components on some phenotypes change as a result of including a moderator in the analysis. Although we cannot comment on all existing gene–environment interactions in this article, we discuss a few of the most replicated effects to better illustrate this definition (for an overview, see Dick, 2011 ; Spinath & Johnson, 2011 ).
The most widely known moderator of the association between genetic and environmental contributions to various traits is age. In general, genetic effects on a trait increase with time, and shared environmental effects decrease; therefore, age is considered to moderate genetic and environmental contributions by “activating” genetic risk as people get older ( Burt, 2011 ). For example, shared environmental effects explained more variance in depression among 8- to 10-year-olds (76%) than among 11- to 17-year-olds (47%). Conversely, additive genetic effects explained less variance in depression among 8- to 10-year-olds (0%) and more among 11- to 17-year-olds (29%; Rice, Harold, & Thaper, 2002 ). Rice et al. (2002) found nonshared environmental influence to be the same across 8- to 10-year-olds and 11- to 17-year-olds (24%). Similarly, age moderates the relationship between genetic and environmental effects and general cognitive ability. Using six twin studies from four countries, Haworth et al. (2009) found that additive genetic effects account for 41% of the variance in general cognitive ability in childhood, 55% in adolescence, and 66% in young adulthood. A similar reversal was found for shared environment estimates: 33% in childhood, 18% in adolescence, and 16% in young adulthood. Similar results have also been found for substance use ( Kendler, Schmitt, Aggen, & Prescott, 2008 ) and peer-group deviance ( Kendler et al., 2007 ).
Evidence of the moderating effect of gender in gene–environment interactions is less clear. For example, regarding tobacco, alcohol, and drug use, Han, McGue, and Iacono (1999) reported higher estimates of additive genetic effects and lower estimates of shared environment for males than for females. However, these differences were not statistically significant. In general, analyzing samples with reported low substance use (females typically use fewer substances than males; younger adolescents typically use fewer substances than older adolescents and adults) usually results in higher estimates of shared environmental effects. Therefore, it seems likely that the shared environment is particularly important in explaining low levels of drug use. It also indicates that genetic effects are more important when the risk of using drugs increases (known as activator effects; Burt, 2011 ).
In terms of depression, Silberg, Rutter, D’Onofrio, and Eaves (2003) reported significantly stronger additive genetic effects for females (and stronger shared environment effects for males). Yet Rice et al. (2002) found the opposite pattern. Kendler, Prescott, Myers, and Neale (2003) did not find any significant difference in shared environment and additive genetic effects between men and women across latent factors representing behavioral disinhibition and internalizing problems. This, along with other evidence, further suggests a limited moderating effect of gender on the relationships between (1) a 2 , c 2 , and e 2 and (2) internalizing and externalizing outcomes ( Eley, Lichtenstein, & Stevenson, 1999 ; Taylor, McGue, & Iacono, 2000).
An additional moderator on male adolescent externalizing appears to be residency in urban versus rural environments ( Rose, Dick, Viken, & Kaprio, 2001 ; Legrand, Keyes, McGue, Iacono, & Krueger, 2008 ). In their replication of Rose et al. (2001) , Legrand et al. (2008) found that genetic effects were stronger for substance use and antisocial behavior among male adolescents living in urban environments ( a 2 ranging from .49 to .57; c 2 ranging from .05 to .22), and that shared environmental effects were stronger for those in rural environments ( a 2 ranging from .02 to .04; c 2 ranging from .35 to .62). In this way, living in a rural environment acts as a deactivator of genetic risk for antisocial behavior, at least for males.
Potentially relating to the moderating effect of urban versus rural environments, SES also appears to moderate the association among variance components and antisocial behavior. For example, Tuvblad, Grann, and Lichtenstein (2006) found a gene–environment interaction for adolescent antisocial behavior as a result of neighborhood socioeconomic conditions. Genetic effects were stronger for male adolescents from higher-SES neighborhoods, and shared environment effects were stronger for male adolescents from lower-SES neighborhoods. Again, however, this was the case only for male adolescents. Specifically, in less advantaged neighborhoods, additive genetic effects were estimated at 1% and shared environment effects at 69%. In more advantaged neighborhoods, additive effects were estimated at 37% and shared environment effects at 13%. It is unclear what other kinds of variables might explain shared environmental effects for female adolescents exhibiting antisocial behavior.
Peer deviance has also been shown to be an activator of genetic risk in several studies (e.g., Agrawal et al., 2010 ; Beaver, Gibson et al., 2009 ; Button, Stallings, Hyun Rhee, Boardman, & Hewitt, 2009 ; Dick et al., 2007 ; Guo, Elder, Cai, & Hamilton, 2009 ; Harden, Hill, Turkheimer, & Emery, 2008 ). For example, using a twin sample of young adult women, Agrawal et al. (2010) found that women who had more friends who used substances had a greater heritability for regular substance involvement. This effect was interpreted within a gene–environment correlation framework: Young women may select friends on the basis of their similar dispositional traits (e.g., being open to using alcohol and drugs); the social environment of those friends then modifies the genetic architecture of substance-use involvement.
Finally, perceptions of parenting and parenting behaviors have acted as moderators of genetic versus environmental risk in several studies on outcomes such as personality and parent–child relationships ( Krueger, South, Johnson, & Iacono, 2008 ; South, Krueger, Johnson, & Iacono 2008 ), parenting and antisocial behavior ( Button et al., 2007 ; Feinberg, Button, Neiderhiser, Reiss, & Hetherington, 2007 ), parental divorce and disordered eating ( Suisman, Burt, McGue, Iacono, & Klump, 2011 ), divorce and child problem behavior (Robbers et al., 2011), and so on. Although we cannot cover this topic extensively in this review, we provide a few examples to illustrate the importance of continued research on parenting using G×E methods (again, for an extensive review, see Dick, 2011 ).
Recall our earlier discussion on the negligible shared environmental effects on personality. Krueger et al. (2008) found that shared environmental factors became increasingly influential when adolescents perceived greater levels of parent–child conflict for the personality trait of negative emotionality, specifically. Moreover, South et al. (2008) found that the genetic versus environmental contributions to parent–child relationship quality varied as a function of their child’s personality. For example, South et al. found that genetic risk in parent involvement increased as a function of children’s general negative emotionality but that shared environmental influences on children’s positive regard for parents increased as a function of children’s negative emotionality. Compared to the main-effects research on personality we discussed earlier, these examples illustrate the complex etiology of parenting in adolescent development and the need for continued examinations of gene–environment interplay.
Conclusion on Quantitative Behavioral Genetics Research
On the basis of these findings of moderators of genetic and environmental effects, we can make several conclusions. First, it is likely that age moderates genetic versus environmental influences on most traits; shared environmental effects diminish with age, whereas genetic effects appear to increase with age. This makes sense given that children spend less time in the shared family environment as they get older and are more likely to seek out environments that match their inherent likes and dislikes (gene–environment correlation; Scarr & McCartney, 1983 ). We can also conclude that some things about SES and rural versus urban living environments appear to have been identified as particularly influential shared environmental effects. This further justifies the need to better understand the relationship between SES and rural versus urban environments in an effort to identify those who are at risk for adjustment problems and to inform interventions. Moreover, we can conclude that family relationship quality, particularly among siblings, may bring an important shared environment contribution to traits such as childhood conduct problems and adolescent substance use. More research is needed overall that seeks to decompose correlations between family outcomes and child behavioral outcomes (particularly those with moderate to substantial shared environmental effects) to better understand what the shared environment really is and how it works. Continued research is needed in on gene–environment interactions to understand when the shared environment is particularly influential. In general, we need more research that seeks to understand gene–environment interplay in the context of development ( Burt, 2011 ; Spinath & Johnson, 2011 ).
To be clear, we and others (e.g., Neiderhiser, 2011 ; Vrieze, Iacono, & McGue, 2012 ) believe that there is much to be learned about the shared environment and gene–environment interplay in the associations between family processes and child and adolescent outcomes. The use of genetically informed samples allows family scientists to fully analyze specific shared environmental variants, and therefore to better understand them. Behavioral genetic analyses to understand gene–environment correlations and interactions are useful statistical tools available to test theories that account for genetic and environmental effects. To perform a behavioral genetics analysis, information about how children are genetically related (e.g., full biological offspring of parents) or unrelated (e.g., adopted, step, blended) to parents and siblings is needed to determine groups. If large, genetically informed samples are not available to family researchers, there are public genetically informed data sets available (e.g., Add Health; Beaver, DeLisi, Wright, & Vaughn, 2009 ; Beaver, Gibson et al., 2009 ). Regardless, it is important to at least consider how study results concerning associations between family processes and child outcomes may be mediated by genetic versus environmental mechanisms, as well as to note how study findings fit with previously published behavioral genetics research.
Switching Gears: A Brief Look at New Methods in Molecular Genetics
Although the focus of this article is on how family researchers can extend quantitative behavioral genetics research by identifying specific shared environment variants and utilizing methods of gene–environment interplay, we believe that it is crucial to briefly review and comment on molecular genetics research because of the increasing number of molecular genetics studies on psychopathology. In fact, it has been argued that the future of understanding genetic and environmental influences depends considerably on the ability to detect specific genetic variants using molecular genetic approaches ( McGue, 2008 ).
Rather than looking at ballpark estimates of genetic effects, new methods are trying to pinpoint specific genetic influences. There are several methods for identifying genetic influences that are beyond the scope of our review. We provide some examples of these methods to begin to familiarize readers with the names and basic ideas involved. One of the earliest examples is linkage analysis for single-gene disorders ( Plomin et al., 2008 ). Single genes have been found to be associated with very rare disorders, such as Huntington’s disease. However, linkage analysis has been less successful in identifying genes for complex disorders (when many genes are involved), which is thought to include most psychological traits ( Plomin et al., 2008 ). Allelic association and candidate gene studies are also commonly published, which have generally been difficult to replicate ( Chabris et al., 2012 ; Tabor, Risch, & Myers, 2002 ).
Here, we focus on genome-wide association studies (GWAS; e.g., International Schizophrenia Consortium, 2009 ; Plomin et al., 2008 ), which are one of the latest developments among rapidly developing human genomic methodologies. For example, the total number of publications of GWAS findings has increased from less than 100 in 2007 to nearly 1,000 in 2011 ( Hindorff et al., 2012 ). Our goal for this section is to provide a brief introduction to GWAS and landmark molecular G×E studies for the unfamiliar reader in family science. For a comprehensive review of genomic methods and issues related to GWAS, we encourage readers to review Vrieze et al. (2012) .
What Is GWAS?
Genome-wide association studies examine the specific genetic influence or effect of what varies within our genes: an allele. Readers may be familiar with the word SNP (single-nucleotide polymorphism) from brief reports on major GWAS findings. Alleles are measured by examining those SNPs. GWAS is a method for examining associations between all or most SNP markers and a phenotype of interest, sometimes in a case-control format ( Corvin, Craddock, & Sullivan, 2010 ). Usually, more than 300,000 SNP markers are examined in association with a phenotype of interest ( Hardy & Singleton, 2009 ).
The unit of analysis, the SNP, is overwhelmingly binary (e.g., paired adenine and guanine, cytosine, and thymine) and is what varies in the human genome. SNPs are collected through DNA assays, which denote a comprehensive but not all-inclusive set of genetic markers across the human genome. Hundreds of thousands of SNPs are compared in association with a phenotype of interest, such as alcoholism. If, after controlling for multiple testing and replication across multiple data sources, cases (e.g., alcoholics) are more likely to have a certain number of alleles on or across a certain number of genes compared to controls (e.g., nonalcoholics), we could conclude that those genetic variants are central in genetically influencing a given trait (e.g., alcoholism). Major benefits of GWAS is that it does not require an a priori hypothesis, and many genetic variants can be analyzed at once ( Hardy & Singleton, 2009 ).
GWAS Assumptions
Our purpose here is not to overwhelm readers with detailed information but to briefly introduce the big picture behind GWAS research (for a detailed review, see Corvin et al., 2010 ).
First, it is necessary to discuss the common disease and common variant hypothesis (CDCV; Hardy & Singleton, 2009 ). This hypothesis predicts that common disease-causing genetic variants are found in all human populations who carry that disease. The opposing model is the common disease and rare variant hypothesis (CDRV). This hypothesis proposes that disease-related genetic variants are found in different human populations who carry the disease. If CDRV proves the case for most diseases, then GWAS will be much less valuable, because there will be different genetic causes for the same disease. Therefore, GWAS operates with the assumption of CDCV because it seeks to identify genetic variants individuals have in common in relation to outcomes.
Related to the difference in these hypotheses is the need to control for genetic ancestry ( McCarthy et al., 2008 ). This helps reduce potential confounds of causal genetic variants across different human populations. Genetic ancestry refers to a person’s shared ancestry (not race or ethnicity) and is determined by geographic location of origin. Racial categories are typically socially constructed and have limited biological utility. Differences exist between genetic ancestries mostly because of chance or evolutionary selection. For example, East Asians are at a higher risk for an ALDH2 (enzyme) deficiency because of an inability to break down acetaldehyde into acetate when drinking alcohol. Therefore, some experience a flushing sensation and an unpleasant response to alcohol. Consequently, people with this deficiency have a reduced likelihood of alcoholism ( Eng, Luczack, & Wall, 2007 ). An examination of these genetic variants in a study (not controlling for genetic ancestry) would reveal an effect for this genetic variant on alcoholism but would fail to recognize that it is significant for only a subpopulation (see Ntzani, Liberopoulos, Manolio, & Ioannidis, 2012 ).
Finally, and as previously mentioned, GWAS requires large samples and replication as a result of restrictions on statistical power ( McCarthy et al., 2008 ). This is because of the simultaneously testing of thousands of regressions (alleles predicting phenotypes) and the need to correct for multiple testing (to control for a Type I error, or false positive). Because of the need for large sample sizes to detect an effect, many studies are involved in consortiums to pool together data for sufficient power.
Major GWAS Findings
Rather than broadly review several findings, we focus on one major research finding that is generally consistent with most GWAS findings on many psychosocial traits. As discussed earlier, quantitative behavioral genetics research has shown large additive genetic effects for schizophrenia and bipolar disorder (approximately 60%; Lichtenstein et al., 2009 ). The International Schizophrenia Consortium (2009) , led by S. M. Purcell, conducted a large-scale GWAS analysis (3,000 cases and matching controls without schizophrenia) and found some evidence of 74,000 SNPs in the Chromosome 6 region. Those same SNPs that predicted schizophrenia also predicted bipolar disorder, thus indicating a potential shared etiology of those diseases. In this study, however, only 3% of the variance in schizophrenia could be explained by these genetic variants alone. However, by using simulation techniques (and making certain assumptions), findings showed the variance explained may be nearly 30%. Still, these estimates are remarkably different from the heritability predicted by twin studies. This example reflects what is reported across various GWAS analyses: very small genetic effects that were previously thought to be quite large, based on quantitative behavioral genetics methods. This has been referred to as the case of the missing heritability ( Maher, 2008 ), and newer methods continue to be developed and refined to help deal with some of the assumptions of GWAS.
Recently, there has been attention to methods such as versatile gene-based tests ( Liu et al., 2010 ), genome-wide complex trait analysis ( Yang, Lee, Goddard, & Visscher, 2011 ), pathway analysis ( Hong, Pawitan, Magnusson, & Prince, 2009 ), and methods that use genetic scoring ( GIANT Consortium, 2010 ; Vrieze et al., 2011 ), which seems especially promising. Genetic scoring methods take the top SNPs from a GWAS (or even better, a meta-analysis of GWAS results), weight them according to their strength of effects, and use them in subsequent analyses. The design and implementation of new methods will clearly continue in the goal of discovering specific genetic influences of various complex traits.
G×E Using Molecular Genetic Methods
There is also a large body of research that utilizes molecular genetics techniques in G×E (for a review, see Burt, 2011 ; Dick, 2011 ; Uher, 2011 ). In these types of analyses, a genetic variant is targeted on the basis of a hypothesis (e.g., the connection between serotonin or dopamine transporters in relation to pharmaceuticals) and is modeled to interact with some environmental factor to predict an outcome. Perhaps the best known of these studies is the research by Caspi and colleagues ( Caspi et al., 2002 ; Caspi et al., 2003 ). In the first molecular genetic G×E report, Caspi et al. (2002) used a candidate gene approach that focused on the MAOA gene, as previous animal and human research has found that the absence of MAOA is associated with aggression in men ( Brunner, Nelen, Breakefield, Ropers, & van Oost, 1993 ; Uher, 2011 ). Caspi et al. found that while the MAOA gene was not associated with antisocial behavior, those with a particular allele frequency on the MAOA gene and who had also experienced child abuse had an increased risk of adult antisocial behavior. In 2003, Caspi et al. found a similar pattern of results utilizing the SERT gene (involved in serotonin regulation), stressful life events, and depression.
These initial findings led to much excitement across a variety of fields. However, replication of Caspi’s work has been somewhat controversial. In their meta-analysis of eight studies, Taylor and Kim-Cohen (2007) found evidence for the MAOA and antisocial behavior interaction, but the stated evidence was preliminary, and more replication was needed. However, Risch et al. (2009) acquired data from 14 independent studies ( N = 14,000+) and found no conclusive evidence of the interaction of SERT, child abuse, and depression. In response, Caspi, Hariri, Holmes, Huher, and Moffitt (2010) reviewed several human observational studies, neuroscience studies, and animal research studies to provide more conclusive evidence on G×E with SERT. Uher (2011) also reviewed potential reasons for the inconclusive findings, including methodological (e.g., use of structured interviews versus self-reports of child abuse) and statistical issues that arise when analyzing categorical outcomes in G×E interactions (for more detail, see Eaves, 2006 ).
This illustrates the mixed and inconclusive findings in the field and underscores the need for continued replication. Particularly when it comes to G×E using molecular methods (and allelic association or candidate gene studies that test main genetic variant effects, for that matter), it is important to find out whether the results replicate before generating too much excitement, too quickly. In fact, the journal Behavior Genetics has required direct replication of any candidate gene association study to be considered for publication ( Hewitt, 2012 ). Journals with reviewers who are not as familiar with this field may be seeing an increasing amount of candidate gene studies submitted for publication (that may fail to replicate; see Duncan & Keller, 2011 ).
Conclusions on New Methods in Molecular Genetics
Overall, GWAS findings have found rather small effects on traits that we think of as largely heritable. Several factors may contribute to these findings. First, the effects themselves may actually be quite small (CDCV hypothesis), or the effects may be rare and differ across populations (CDRV hypothesis). Second, correlation is not causation; just because the SNP and phenotype are highly correlated does not mean that the SNP is causing variation in the phenotype. The identified allele may not be the causal mechanism, but rather may be just correlated with the causal genetic variant (called linkage disequilibrium). Third, the small effects may be due to an interaction between alleles and the environment (which, again, are often hard to replicate). Fourth, genetic effects may be confounded if results are not controlled for genetic ancestry. Finally, alleles may not be the appropriate level of analysis for discovering genetic variant effects—hence the need for additional methods such as versatile gene-based tests ( Liu et al., 2010 ), genome-wide complex trait analysis ( Yang et al., 2011 ), path analysis ( Hong et al., 2009 ), and genetic scoring ( GIANT Consortium, 2010 ; Vrieze et al., 2011 ). Moreover, research that utilizes G×E via molecular genetics methods such as candidate gene studies will continue to grow, but caution is warranted in interpreting nonreplicated findings.
Molecular genetics methods and findings on complex traits such as psychopathology are only beginning to emerge. Even though only very small effects have been detected using GWAS, this research could still potentially have a large payoff. For example, findings may be helpful in understanding the etiology of mental health disorders that profoundly affect family systems (e.g. schizophrenia and bipolar disorder; International Schizophrenia Consortium, 2009 ). Family practitioners may someday utilize GWAS or other molecular genetics findings to inform clinical interventions with families that promote healthy family functioning. Such interventions may be achieved through psychoeducation and discussion of research findings with families. At this point, however, we are not there yet, and we have a long way to go.
Conclusions on Behavioral and Molecular Genetics for Family Researchers
In summary, we believe that understanding behavioral and molecular genetics research can substantially expand our understanding of families. Moreover, through collaboration across family and behavior genetics disciplines, we can help inform a broader, more complete understanding of the complex etiology of family health and well-being. Members of our field are particularly well placed to contribute to the understanding of shared environmental effects and the impact of gene–environment interplay. The main point is this: If genes and the environment are both having effects on human development (and most of our theories tell us so), then our odds of understanding the environmental influences are better if we can understand and systematically rule out genetic influences.
Acknowledgments
Diana R. Samek was supported by Grant No. MH017069 from the National Institute of Mental Health postdoctoral training grant.
- Agrawal A, Balasubramanian S, Smith EK, Madden PAF, Bucholz KK, Heath AC, Lynskey MT. Peer substance involvement modifies genetic influences on regular substance involvement in young women. Addiction. 2010; 105 :1844–1853. doi:10.1111/j.1360-0443.2010.02993.x. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Bazzett TJ. An introduction to behavior genetics. Sinauer; Sunderland, MA: 2008. [ Google Scholar ]
- Beaver KM, DeLisi M, Wright JP, Vaughn MG. Gene–environment interplay and delinquent involvement: Evidence of direct, indirect, and interactive effects. Journal of Adolescent Research. 2009; 24 :147–168. doi:10.1177/0743558408329952. [ Google Scholar ]
- Beaver KM, Gibson CL, Turner MG, DeLisi M, Vaughn MG, Holand A. Stability of peer associations: A biosocial test of Warr’s stick-friends hypothesis. Crime and Delinquency. 2009; 57 :907–926. doi:10.1177/0011128709332660. [ Google Scholar ]
- Behavior Genetics Association (n.d.). Retrieved from http://www.bga.org/ [ Google Scholar ]
- Borkenau P, Riemann R, Angleitner A, Spinath FM. Similarity of childhood experiences and personality resemblance in monozygotic and dizygotic twins: A test of the equal environments assumption. Personality and Individual Differences. 2002; 33 :261–269. [ Google Scholar ]
- Bouchard TJ, McGue M. Genetic and environmental influences on human psychological differences. Journal of Neurobiology. 2003; 54 :4–45. doi:10.1002/neu.10160. [ PubMed ] [ Google Scholar ]
- Brunner HG, Nelen M, Breakefield XO, Ropers HH, van Oost BA. Abnormal behavior associated with a point mutation in the structural gene for monoamine oxidase A. Science. 1993; 262 :578–580. [ PubMed ] [ Google Scholar ]
- Buchanan JP, McGue M, Keyes M, Iacono WG. Are there shared environment influences on adolescent behavior? Evidence from a study of adoptive siblings. Behavior Genetics. 2009; 39 :532–540. doi:10.1007/s10519-009-9283-y. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Bulik CM, Sullivan PF, Wade TD, Kendler KS. Twin studies of eating disorders: A review. International Journal of Eating Disorders. 2000; 27 :1–20. [ PubMed ] [ Google Scholar ]
- Burt A. Some key issues in the study of gene–environment interplay: Activation, deactivation, and the role of development. Research in Human Development. 2011; 8 :192–210. doi:10.1080/15427609.2011.625323. [ Google Scholar ]
- Burt SA. Rethinking environmental contributions to child and adolescent psychopathology: A meta-analysis of shared environmental influences. Psychological Bulletin. 2009; 135 :608–637. doi:10.1037/a0015702. [ PubMed ] [ Google Scholar ]
- Burt SA, Klahr AM, Rueter MA, McGue M, Iacono WG. Confirming the etiology of adolescent acting-out behaviors: An examination of observer-ratings in a sample of adoptive and biological siblings. Journal of Child Psychology and Psychiatry. 2011; 52 :519–526. doi:10.1111/j.1469-7610.2010.02334.x. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Burt SA, McGue M, Iacono WG, Krueger RF. Differential parent–child relationships and adolescent externalizing symptoms: Cross-lagged analyses with a monozygotic twin differences design. Developmental Psychology. 2006; 42 :1289–1298. doi:10.1037/0012-1649.42.6.1289. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Burt SA, McGue M, Krueger RF, Iacono WG. Environmental contributions to adolescent delinquency: A fresh look at the shared environment. Journal of Abnormal Child Psychology. 2007; 35 :787–800. doi:10.1007/s10802-007-9135-2. [ PubMed ] [ Google Scholar ]
- Bussell DA, Neiderhiser JM, Pike A, Plomin R, Simmens S, Howe GW, Reiss D. Adolescents’ relationships to siblings and mothers: A multivariate genetic analysis. Developmental Psychology. 1999; 35 :1248–1259. [ PubMed ] [ Google Scholar ]
- Button TM, Corley RP, Rhee SH, Hewitt JK, Young SE, Stallings MC. Delinquent peer affiliation and conduct problems: A twin study. Journal of Abnormal Psychology. 2007; 116 :554–64. doi:10.1037/0021-843X.116.3.554. [ PubMed ] [ Google Scholar ]
- Button TMM, Stallings MC, Hyun Rhee S, Corley RP, Boardman JD, Hewitt JK. Perceived peer delinquency and the genetic predisposition for substance dependence vulnerability. Drug and Alcohol Dependence. 2009; 100 :1–8. doi:10.1016/j.drugalcdep.2008.08.014. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Caspi A, Hariri AR, Holmes A, Uher R, Moffitt TE. Genetic sensitivity to the environment: The case of the serotonin transporter gene and its implications for studying complex diseases and traits. American Journal of Psychiatry. 2010; 167 :509–527. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Caspi A, McClay J, Moffitt TE, Mill J, Martin J, Craig IW, Poulton R. Role of genotype in the cycle of violence in maltreated children. Science. 2002; 297 :851–857. [ PubMed ] [ Google Scholar ]
- Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H, Poulton R. Influence of life stress on depression: Moderation by a polymorphism in the 5-HTT gene. Science. 2003; 301 :386–389. [ PubMed ] [ Google Scholar ]
- Chabris CF, Hebert BM, Benjamin DJ, Beauchamp J, Cesarini D, van der Loos M, Laibson D. Most reported genetic associations with general intelligence are probably false positives. Psychological Science. 2012; 23 :1314–1323. doi:10.1177/0956797611435528. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Collins WA, Maccoby EE, Steinberg L, Hetherington EM, Bornstein MH. Contemporary research on parenting: The case for nature and nurture. American Psychologist. 2000; 55 :218–232. doi:10.1037/0003-066X.55.2.218. [ PubMed ] [ Google Scholar ]
- Corvin A, Craddock N, Sullivan PF. Genome-wide association studies: A primer. Psychological Medicine. 2010; 40 :1063–1077. doi:10.1017/S0033291709991723. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Dar-Nimrod I, Heine SJ. Genetic essentialism: On the deceptive determinism of DNA. Psychological Bulletin. 2011; 137 :800–818. doi:10.1037/a0021860. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Deater-Deckard K. Parenting and child behavioral adjustment in early childhood: A quantitative genetic approach to studying family processes. Child Development. 2000; 71 :468–484. doi:10.1111/1467-8624.00158. [ PubMed ] [ Google Scholar ]
- Dick DM. Gene–environment interaction in psychological traits and disorders. Annual Review of Clinical Psychology. 2011; 7 :783–409. doi:10.1146/annurev-clinpsy-032210-104518. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Dick DM, Latendresse SJ, Riley B. Incorporating genetics into your studies: A guide for social scientists. Frontiers in Psychiatry. 2011; 2 :1–11. doi:10.3389/fpsyt.2011.00017. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Dick DM, Pagan JL, Viken R, Purcell S, Kaprio J, Pulkkinen L, Rose RJ. Changing environmental influences on substance use across development. Twin Research on Human Genetics. 2007; 10 :315–326. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- DiLalla LF. Behavior genetics of aggression in children: Review and future directions. Developmental Review. 2002; 22 :593–622. doi:10.1016/S0273-2297(02)00504-X. [ Google Scholar ]
- D’Onofrio B. Children of twins design. Encyclopedia of Statistics in Behavioral Science. 2005 doi:10.1002/0470013192.bsa088. [ Google Scholar ]
- D’Onofrio BM, Lahey BB. Biosocial influences on the family: A decade review. Journal of Marriage and Family. 2010; 72 :762–782. doi:10.1111/j.1741-3737.2010.00729.x. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Duncan LE, Keller MC. A critical review of the first 10 years of gene-by-environment interaction research in psychiatry. American Journal of Psychiatry. 2011; 168 :1041–1049. doi:10.1176/appi.ajp.2011.11020191. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Duyme M, Dumaret A, Tomkiewicz S. How can we boost IQs of “dull children”? A late adoption study. Proceedings of the National Academy of Sciences of the United States of America. 1999; 96 :8790–8794. doi:10.1073/pnas.96.15.8790. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Eaves LJ. Genotype × environment interaction in psychopathology: Fact or artifact? Twin Research and Human Genetics. 2006; 9 :1–8. [ PubMed ] [ Google Scholar ]
- Eley TC, Lichtenstein P, Stevenson J. Sex differences in the etiology of aggressive and nonaggressive antisocial behavior from two twin studies. Child Development. 1999; 70 :155–168. doi:10.1111/1467-8624.00012. [ PubMed ] [ Google Scholar ]
- Elkins IJ, McGue M, Iacono WG. Genetic and environmental influences on parent–son relationships: Evidence for increasing genetic influence during adolescence. Developmental Psychology. 1997; 33 :351–363. doi:10.1037//0012-1649.33.2.351. [ PubMed ] [ Google Scholar ]
- Eng MY, Luczack SE, Wall TL. ALDH2, ADH1B, and ADH1C genotypes in Asians: A literature review. Alcohol Research and Health. 2007; 30 :22–27. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Fagan AA, Najman JM. The relative contributions of parent and sibling substance use to adolescent tobacco, alcohol, and other drug use. Journal of Drug Issues. 2005; 35 :869–884. doi:10.1207/S15328007SEM0803_5. [ Google Scholar ]
- Feinberg ME, Button TM, Neiderhiser JM, Reiss D, Hetherington EM. Parenting and adolescent antisocial behavior and depression: Evidence of genotype × parenting environment interaction. Archives of General Psychiatry. 2007; 64 :457–465. [ PubMed ] [ Google Scholar ]
- Feinberg M, Neiderhiser J, Howe G, Hetherington E. Adolescent, parent, and observer perceptions of parenting: Genetic and environmental influences on shared and distinct perceptions. Child Development. 2001; 72 :1266–1284. doi:10.1111/1467-8624.00346. [ PubMed ] [ Google Scholar ]
- GIANT (Genetic Investigation of Anthropocentric Traits) Consortium Hundreds of variants clustered in genomic loci and biological pathways affect human height. Nature. 2010; 467 :832–838. doi:10.1038/nature09410. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Guo G, Elder GH, Cai T, Hamilton N. Gene–environment interactions: Peers’ alcohol use moderates genetic contribution to adolescent drinking behavior. Social Science Research. 2009; 38 :213–224. doi:10.1016/j.ssresearch.2008.04.002. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Hamilton MB. Population genetics. Blackwell; West Sussex, UK: 2009. [ Google Scholar ]
- Han C, McGue MK, Iacono WG. Lifetime tobacco, alcohol and other substance use in adolescent Minnesota twins: Univariate and multivariate behavioral genetic analyses. Addiction. 1999; 94 :981–993. doi:10.1046/j.1360-0443.1999.9479814.x. [ PubMed ] [ Google Scholar ]
- Harden KP, Hill JE, Turkheimer E, Emery RE. Gene–environment correlation and interaction in peer effects on adolescent alcohol and tobacco use. Behavior Genetics. 2008; 38 :339–347. doi:10.1007/s10519-008-9202-7. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Hardy J, Singleton A. Genomewide association studies and human disease. New England Journal of Medicine. 2009; 36 :1759–1768. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Hatemi PK, Dawes CT, Frost-Keller A, Settle JE, Verhulst B. Integrating social science and genetics: News from the political front. Biodemography and Social Biology. 2011; 57 :67–87. doi:10.1080/19485565.2011.568276. [ PubMed ] [ Google Scholar ]
- Haworth CMA, Wright MJ, Luciano M, Martin NG, de Geus EJC, van Beijsterveldt CEM, Plomin R. The heritability of general cognitive ability increases linearly from childhood to young adulthood. Molecular Psychiatry. 2009; 15 :1112–1120. doi:10.1038/mp.2009.55. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Hewitt JK. Editorial policy on candidate gene association and candidate gene-by-environment interaction studies of complex traits. Behavior Genetics. 2012; 42 :1–2. doi:10.1007/s10519-011-9504-z. [ PubMed ] [ Google Scholar ]
- Hindorff LA, MacArthur J, Wise A, Junkins HA, Hall PN, Klemm AK, Manolio TA. A catalog of published genome-wide association studies. 2012 Retrieved from http://www.genome.gov/gwastudies . [ Google Scholar ]
- Hong M, Pawitan Y, Magnusson PKE, Prince J. Strategies and issues in the detection of pathway enrichment in genome-wide association studies. Human Genetics. 2009; 126 :289–301. doi:10.1007/s00439-009-0676-z. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Hopfer CJ, Crowley TJ, Hewitt JK. Review of twin and adoption studies of adolescent substance use. Journal of the American Academy of Child and Adolescent Psychiatry. 2003; 43 :710–719. doi:10.1097/01.CHI.0000046848.56865.54. [ PubMed ] [ Google Scholar ]
- Horwitz BN, Neiderhiser JM. Gene–environment interplay, family relationships, and child adjustment. Journal of Marriage and Family. 2011; 73 :804–816. doi:10.1111/j.1741-3737.2011.00846.x. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Iacono WG, McGue M. Minnesota Twin Family Study. Twin Research. 2002; 5 :482–487. [ PubMed ] [ Google Scholar ]
- International Schizophrenia Consortium. Purcell SM, Wray NR, Stone JL, Visscher PM, O’Donovan MC, Sklar P. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature. 2009; 460 :475–752. doi:10.1038/nature08185. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Jang KL, Livesley WJ, Vernon PA. Heritability of the big five personality dimensions and their facets: A twin study. Journal of Personality. 1996; 64 :577–591. [ PubMed ] [ Google Scholar ]
- Kendler KS, Gardner CO., Jr. Twin studies of adult psychiatric and substance dependence disorders: Are they biased by differences in the environmental experiences of monozygotic and dizygotic twins in childhood and adolescence? Psychological Medicine. 1998; 28 :625–633. doi:10.1017/S0033291798006643. [ PubMed ] [ Google Scholar ]
- Kendler KS, Jacobson KC, Gardner CO, Gillespie N, Aggen SA, Prescott CA. Creating a social world: A developmental twin study of peer-group deviance. Archives of General Psychiatry. 2007; 64 :958–965. doi:10.1001/archpsyc.64.8.958. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Kendler KS, Prescott CA. Genes, environment and psychopathology: Understanding the causes of psychiatric and substance use disorders. Guilford Press; Boston, MA: 2006. [ Google Scholar ]
- Kendler KS, Prescott CA, Myers J, Neale MC. The structure of genetic and environmental risk factors for common psychiatric and substance use disorders in men and women. Archives of General Psychiatry. 2003; 60 :929–937. doi:10.1001/archpsyc.60.9.929. [ PubMed ] [ Google Scholar ]
- Kendler KS, Schmitt E, Aggen SH, Prescott CA. Genetic and environmental influences on alcohol, caffeine, cannabis, and nicotine use from early adolescence to middle adulthood. Archives of General Psychiatry. 2008; 65 :674–682. doi:10.1001/archpsyc.65.6.674. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Kim Y, editor. Handbook of behavior genetics. Springer; New York, NY: 2009. [ Google Scholar ]
- Klahr AM, McGue M, Iacono WG, Burt SA. The association between parent–child conflict and adolescent conduct problems over time: Results from a longitudinal adoption study. Journal of Abnormal Psychology. 2011; 120 :46–56. doi:10.1037/a0021350. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Krueger RF, South S, Johnson W, Iacono W. The heritability of personality is not always 50%: Gene–environment interactions and correlations between personality and parenting. Journal of Personality. 2008; 76 :1485–1521. doi:10.1111/j.1467-6494.2008.00529.x. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Latendresse SJ, Rose RJ, Viken RJ, Pulkkinen L, Kaprio J, Dick DM. Examining the etiology of associations between perceived parenting and adolescents’ alcohol use: Common genetic and/or environmental liabilities? Journal of the Study on Alcohol and Drugs. 2010; 71 :313–325. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Legrand LN, Keyes M, McGue M, Iacono WG, Krueger RF. Rural environments reduce the genetic influence on adolescent substance use and rule-breaking behavior. Psychological Medicine. 2008; 38 :1341–1350. doi:10.1017/S0033291707001596. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Leve LD, Neiderhiser JM, Scaramella LV, Reiss D. The early growth and development study: Using the prospective adoption design to examine genotype–environment interplay. Acta Psychologica Sinica. 2008; 10 :1106–1115. doi:10.3724/SP.J.1041.2008.01106. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Lichtenstein P, Yip BH, Bjork C, Pawitan Y, Cannon TD, Sullivan PF, Hultman CM. Common genetic determinants of schizophrenia and bipolar disorder in Swedish families: A population-based study. Lancet. 2009; 373 :234–239. doi:10.1016/S0140-6736(09)60072-6. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Liu JZ, McCrae AF, Nyholt DR, Medland SE, Wray NR, Brown KM, Macgregor S. A versatile gene-based test for genome-wide association studies. American Journal of Human Genetics. 2010; 87 :139–145. doi:10.1016/j.ajhg.2010.06.009. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Loehlin JC. Environment and the behavior genetics of personality: Let me count the ways. Personality and Individual Differences. 2010; 49 :302–305. doi:10.106/j.paid.2009.10.035. [ Google Scholar ]
- Loehlin JC, McCrae RR, Costa PT, Jr., John OP. Heritabilities of common and measure-specific components of the big five personality factors. Journal of Research in Personality. 1998; 32 :431–453. [ Google Scholar ]
- Loehlin JC, Nichols RC. Heredity, environment, and personality. University of Texas Press; Austin, TX: 1976. [ Google Scholar ]
- Lynskey MT, Agrawal A, Health A. Genetically informed research on adolescent substance use: Methods, findings, and challenges. Journal of the American Academy of Child and Adolescent Psychiatry. 2010; 49 :1202–1214. doi:10.1016/j.jaac.2010.09.004. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Maher B. The case of the missing heritability. Nature. 2008; 456 :18–21. [ PubMed ] [ Google Scholar ]
- McCarthy MI, Abecasis G, Cardon LR, Goldstein DB, Little J, Ionnaids JPA, Hirschorn JN. Genome-wide association studies for complex traits: Consensus, uncertainty and challenges. Nature. 2008; 9 :356–369. doi:10.1038/nrg2344. [ PubMed ] [ Google Scholar ]
- McGue M. The end of behavioral genetics? Acta Psychologica Sinica. 2008; 40 :1073–1087. doi:10.3724/SP.J.1041.2008.01073. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- McGue M, Bouchard TJ, Iacono WG, Lykken DT. Behavioral genetics of cognitive ability: A life-span perspective. In: Plomin R, MeClearn GE, editors. Nature, nurture, and psychology. American Psychological Association; Washington, DC: 1993. pp. 59–76. [ Google Scholar ]
- McGue M, Elkins E, Iacono WG. Genetic and environmental influences on adolescent substance use and abuse. American Journal of Medical Genetics (Neuropsychiatric Genetics) 2000; 96 :671–677. [ PubMed ] [ Google Scholar ]
- McGue M, Elkins E, Walden B, Iacono WG. Perceptions of the parent–adolescent relationship: A longitudinal investigation. Developmental Psychology. 2005; 41 :971–984. doi:10.1037/0012-1649.41.6.971. [ PubMed ] [ Google Scholar ]
- McGue M, Iacono WG. Siblings and the socialization of adolescent deviance: An adoption study approach. In: McCartney K, Weinberg R, editors. Experience and development: A festschrift to honor Sandra W. Scarr. Taylor and Francis; London, UK: 2009. pp. 179–201. [ Google Scholar ]
- McGue M, Keyes M, Sharma A, Elkins I, Legrand L, Johnson W, Iacono WG. The environments of adopted and non-adopted youth: Evidence on the range restriction from the Sibling Interaction and Behavior Study (SIBS) Behavior Genetics. 2007; 37 :449–462. doi:10.1007/s10519-007-9142-7. [ PubMed ] [ Google Scholar ]
- McGue M, Sharma A, Benson P. The effect of common rearing on adolescent adjustment: Evidence from a US adoption cohort. Developmental Psychology. 1996; 32 :604–613. doi:00I2-I649/96. [ Google Scholar ]
- Moffitt T, Caspi A, Rutter M. Measured gene–environment interactions in psychopathology: Concepts, research strategies, and implications for research, intervention, and public understanding of genetics. Perspectives on Psychological Science. 2005; 1 [ PubMed ] [ Google Scholar ]
- Neale MC. Biometric models in behavioral genetics. In: Kim Y, editor. Handbook of behavior genetics. Springer; New York, NY: 2009. pp. 15–19. [ Google Scholar ]
- Neiderhiser JM. Gene–environment interplay helps to explain influences of family relationships on adolescent adjustment and development. In: Booth A, McHale SM, Landale NS, editors. Biosocial foundations of family processes. Springer; New York, NY: 2011. pp. 71–84. [ Google Scholar ]
- Neiderhiser JM, Pike A, Hetherington EM, Reiss D. Adolescent perceptions as mediators of parenting: Genetic and environmental contributions. Developmental Psychology. 1998; 34 :1459–1469. doi:10.1037//0012-1649.34.6.1459. [ PubMed ] [ Google Scholar ]
- Neiderhiser JM, Reiss D, Hetherington EM. The nonshared environment in adolescent development (NEAD) project: A longitudinal family study of twins and siblings from adolescence to young adulthood. Twin Research and Human Genetics. 2007; 10 :74–83. doi:10.1375/twin.10.1.74. [ PubMed ] [ Google Scholar ]
- Ntzani EE, Liberopoulos G, Manolio TA, Ioannidis JPA. Consistency of genome-wide associations across major ancestral groups. Human Genetics. 2012; 131 :1057–1071. doi:10.1007/s00439-011-1124-4. [ PubMed ] [ Google Scholar ]
- Pike A, McGuire S, Hetherington EM, Reiss D, Plomin R. Family environment and adolescent depressive symptoms and antisocial behavior: A multivariate genetic analysis. Developmental Psychology. 1996; 32 :590–603. doi:10.1037/0012-1649.32.4.590.l. [ Google Scholar ]
- Plomin R. Commentary: Why are children in the same family so different? Non-shared environment three decades later. International Journal of Epidemiology. 2011; 40 :582–592. doi:10.1093/ije/dyq144. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Plomin R, Daniels D. Why are children in the same family so different from one another? Behavioral and Brain Sciences. 1987; 10 :1–22. [ Google Scholar ]
- Plomin R, DeFries JC, McClearn GE, McGuffin P. Behavior genetics. 4th ed. Worth; New York, NY: 2008. [ Google Scholar ]
- Plomin R, Haworth CMA, Davis OSP. Common disorders are quantitative traits. Nature Reviews Genetics. 2009; 10 :872–872. doi:10.1038/nrg2670. [ PubMed ] [ Google Scholar ]
- Plomin R, Owen MJ, McGuffin P. The genetic basis of complex human behaviors. Science. 1994; 264 :1733–1739. doi:10.1126/science.8209254. [ PubMed ] [ Google Scholar ]
- Plomin R, Rende R. Human behavioral genetics. Annual Review of Psychology. 1990; 42 :161–190. [ PubMed ] [ Google Scholar ]
- Purcell SM. Variance components models for gene-environment interaction in twin analysis. Twin Research. 2002; 5 :554–571. doi:10.1375/136905202762342026. [ PubMed ] [ Google Scholar ]
- Rende R, Slomkowski C. Incorporating the family as a critical context in genetic studies of children: Implications for understanding pathways to risky behavior and substance use. Journal of Pediatric Psychology. 2009; 34 :606–616. doi:10.1093/jpepsy/jsn053. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Rice F, Harold GT, Thapar A. Assessing the effects of age, sex, and shared environment on the genetic aetiology of depression in childhood and adolescence. Journal of Child Psychology and Psychiatry and Allied Disciplines. 2002; 43 :1039–1051. doi:10.1111/1469-7610.00231. [ PubMed ] [ Google Scholar ]
- Risch N, Herrell R, Lehner T, Liang KY, Eaves L, Hoh J, Merikangas KR. Interaction between the serotonin transporter gene (5-HTTLPR), stressful life events, and risk of depression: A meta-analysis. Journal of the American Medical Association. 2009; 301 :2462–2471. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Robbers S, van Oort F, Huizink A, Verhulst F, van Beijsterveldt C, Boomsma D, Bartels M. Childhood problem behavior and parental divorce: Evidence for gene–environment interaction. Social Psychiatry and Psychiatric Epidemiology. 2012; 47 :1539–1548. doi:10.1007/s00127-011-0470-9. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Rose RJ, Dick DM, Viken RJ, Kaprio J. Gene–environment interaction in patterns of adolescent drinking: regional residency moderates longitudinal influences on alcohol use. Alcoholism and Clinical and Experimental Research. 2001; 25 :637–43. [ PubMed ] [ Google Scholar ]
- Rushton JP, Bons TA, Hur Y. The genetics and evolution of the general factor of personality. Journal of Research in Personality. 2008; 42 :1173–1185. doi:10.1016/j.jrp.2008.03.002. [ Google Scholar ]
- Samek DR, Rueter MA. Considerations of elder sibling closeness in predicting younger sibling substance use: Social learning versus social bonding explanations. Journal of Family Psychology. 2011; 25 :931–941. doi:10.1037/a0025857. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Scarr S, McCartney K. How people make their own environments: A theory of genotype–environment effects. Child Development. 1983; 54 :424–435. [ PubMed ] [ Google Scholar ]
- Shelton KH, Harold GT, Fowler TA, Rice FJ, Neale MC, Thapar A, van den Bree MBM. Parent–child relations, conduct problems, and cigarette use in adolescence: Examining the role of genetic and environmental factors on patterns of behavior. Journal of Youth and Adolescence. 2008; 37 :1216–1228. doi:10.1007/s10964-007-9254-7. [ Google Scholar ]
- Silberg J, Rutter M, D’Onofrio B, Eaves L. Genetic and environmental risk factors in adolescent substance use. Journal of Child Psychology and Psychiatry and Allied Disciplines. 2003; 44 :664–676. doi:10.1111/1469-7610.00153. [ PubMed ] [ Google Scholar ]
- Slomkowski C, Rende R, Novak S, Lloyd-Richardson E, Niaura R. Sibling effects on smoking in adolescence: Evidence for social influence from a genetically informative design. Addiction. 2005; 100 :430–438. doi:10.1111/j.1360-0443.2004.00965.x. [ PubMed ] [ Google Scholar ]
- South SC, Krueger RF, Johnson W, Iacono WG. Adolescent personality moderates genetic and environmental influences on relationships with parents. Journal of Personality and Social Psychology. 2008; 94 :899–912. doi:10.1037/0022-3514.94.5.899. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Spinath FM, Johnson W. Behavior genetics. In: Chamorro-Premusic T, von Stumm S, Furnham A, editors. The Wiley-Blackwell handbook of individual differences. Blackwell; West Sussex, UK: 2011. pp. 271–304. [ Google Scholar ]
- Stoolmiller M. Implications of the restricted range of family environments for estimates of heritability and nonshared environment in behavior-genetic adoption studies. Psychological Bulletin. 1999; 125 :392–409. doi:10.1037/0033-2909.125.4.392. [ PubMed ] [ Google Scholar ]
- Suisman JL, Burt SA, McGue M, Iacono WG, Klump KL. Parental divorce and disordered eating: An investigation of gene–environment interaction. International Journal of Eating Disorders. 2011; 44 :169–177. doi:10.1002/eat.20866. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Tabor HK, Risch NJ, Myers RM. Candidate-gene approaches for studying complex genetic traits: Practical considerations. Nature Reviews Genetics. 2002; 3 :391–397. [ PubMed ] [ Google Scholar ]
- Taylor A, Kim-Cohen J. Meta-analysis of gene-environment interactions in developmental psychopathology. Developmental Psychopathology. 2007; 19 :1029–1037. [ PubMed ] [ Google Scholar ]
- Taylor J, McGue M, Iacono W. Sex differences, assortative mating, and cultural transmission effects on adolescent delinquency: A twin family study. Journal of Child Psychology and Psychiatry and Allied Disciplines. 2003; 41 :443–440. doi:10.1017/S0021963000005692. [ PubMed ] [ Google Scholar ]
- Tellegen A, Lykken DT, Bouchard TJ, Jr., Wilcox KJ, Segal NL, Rich S. Personality similarity in twins reared apart and together. Journal of Personality and Social Psychology. 1988; 54 :1031–1039. doi:10.1037/0022-3514.54.6.1031. [ PubMed ] [ Google Scholar ]
- Turkheimer E. Genetics and human agency: Comment on Dar-Nimrod and Heine (2011) Psychological Bulletin. 2011; 137 :825–828. doi:10.1037/a0024306. [ PubMed ] [ Google Scholar ]
- Turkheimer E, Waldron M. Nonshared environment: A theoretical, methodological, and quantitative review. Psychological Bulletin. 2000; 126 :78–108. [ PubMed ] [ Google Scholar ]
- Tuvblad C, Grann M, Lichtenstein P. Heritability for adolescent antisocial behavior: Gene–environment interaction. Journal of Child Psychology and Psychiatry. 2006; 47 :734–743. doi:10.1111/j.1469-7610.2005.01552.x. [ PubMed ] [ Google Scholar ]
- Uher R. Gene–environment interactions. In: Kendler KS, Jaffee SR, Romer D, editors. The dynamic genome and mental health. Oxford University Press; New York, NY: 2011. pp. 28–59. [ Google Scholar ]
- Visscher PM, Hill WG, Wray NR. Heritability in the genomics era: Concepts and misconceptions. Nature Review Genetics. 2008; 9 :255–266. doi:10.1038/nrg2322. [ PubMed ] [ Google Scholar ]
- Visscher PM, Medland SE, Ferreira MAR, Morley KI, Zhu G, Cornes BK, Martin NG. Assumption-free estimation of heritability from genome-wide identity-by-descent sharing between full siblings. PLoS Genetics. 2006; 2 :316–325. doi:10.1371/journal.pgen.0020041.eor. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Vrieze SI, Iacono WG, McGue M. Confluence of genes, environment, development and behavior in a post-GWAS world. Development and Psychopathology. 2012; 24 :1195–1214. doi:10.1017/S0954579412000648. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Vrieze SI, McGue M, Miller MB, Legrand LN, Schork NJ, Iacono WG. An assessment of the individual and collective effects of variants on height using twins and a developmentally informative study design. PLoS Genetics. 2011; 7 :1–10. doi:10.1371/journal.pgen.1002413. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Yamagata S, Suzuki A, Ando J, Ono Y, Kijima N, Yoshimura K, Jang KL. Is the genetic structure of human personality universal? A cross-cultural twin study from North America, Europe, and Asia. Journal of Personality and Social Psychology. 2006; 90 :987–998. doi:10.1037/0022-3514.90.6.987. [ PubMed ] [ Google Scholar ]
- Yang J, Lee SH, Goddard ME, Visscher PM. GCTA: A tool for genome-wide complex trait analyses. American Journal of Human Genetics. 2011; 1 :76–82. doi:10.1016/j.ajhg.2010.11.011. [ PMC free article ] [ PubMed ] [ Google Scholar ]
This paper is in the following e-collection/theme issue:
Published on 14.8.2024 in Vol 26 (2024)
Impact of an Online Discussion Forum on Self-Guided Internet-Delivered Cognitive Behavioral Therapy for Public Safety Personnel: Randomized Trial
Authors of this article:
Original Paper
- Hugh C McCall 1, 2 , MA ;
- Heather D Hadjistavropoulos 1, 2 , PhD
1 Department of Psychology, University of Regina, Regina, SK, Canada
2 PSPNET, University of Regina, Regina, SK, Canada
Corresponding Author:
Heather D Hadjistavropoulos, PhD
Department of Psychology
University of Regina
3737 Wascana Pkwy
Regina, SK, S4S 0A2
Phone: 1 306 585 5133
Email: [email protected]
Background: Internet-delivered cognitive behavioral therapy (ICBT) is an effective and accessible treatment for various mental health concerns. ICBT has shown promising treatment outcomes among public safety personnel (PSP), who experience high rates of mental health problems and face barriers to accessing other mental health services. Client engagement and clinical outcomes are better in ICBT with therapist guidance, but ICBT is easier to implement on a large scale when it is self-guided. Therefore, it is important to identify strategies to improve outcomes and engagement in self-guided ICBT and other self-guided digital mental health interventions. One such strategy is the use of online discussion forums to provide ICBT clients with opportunities for mutual social support. Self-guided interventions accompanied by online discussion forums have shown excellent treatment outcomes, but there is a need for research experimentally testing the impact of online discussion forums in ICBT.
Objective: We aimed to evaluate a transdiagnostic, self-guided ICBT intervention tailored specifically for PSP (which had not previously been assessed), assess the impact of adding a therapist-moderated online discussion forum on outcomes, and analyze participants’ feedback to inform future research and implementation efforts.
Methods: In this randomized trial, we randomly assigned participating PSP (N=107) to access an 8-week transdiagnostic, self-guided ICBT course with or without a built-in online discussion forum. Enrollment and participation were entirely web-based. We assessed changes in depression, anxiety, and posttraumatic stress as well as several secondary outcome measures (eg, treatment engagement and satisfaction) using questionnaires at the pre-enrollment, 8-week postenrollment, and 20-week postenrollment time points. Mixed methods analyses included multilevel modeling and qualitative content analysis.
Results: Participants engaged minimally with the forum, creating 9 posts. There were no differences in treatment outcomes between participants who were randomly assigned to access the forum (56/107, 52.3%) and those who were not (51/107, 47.7%). Across conditions, participants who reported clinically significant symptoms during enrollment showed large and statistically significant reductions in symptoms ( P <.05 and d >0.97 in all cases). Participants also showed good treatment engagement and satisfaction, with 43% (46/107) of participants fully completing the intervention during the course of the study and 96% (79/82) indicating that the intervention was worth their time.
Conclusions: Previous research has shown excellent clinical outcomes for self-guided ICBT accompanied by discussion forums and good engagement with those forums. Although clinical outcomes in our study were excellent across conditions, engagement with the forum was poor, in contrast to previous research. We discuss several possible interpretations of this finding (eg, related to the population under study or the design of the forum). Our findings highlight a need for more research evaluating the impact of online discussion forums and other strategies for improving outcomes and engagement in self-guided ICBT and other digital mental health interventions.
Trial Registration: ClinicalTrials.gov NCT05145582; https://clinicaltrials.gov/study/NCT05145582
Introduction
Internet-delivered cognitive behavioral therapy (ICBT) is a psychological treatment in which clients learn evidence-based cognitive behavioral therapy treatment strategies via web-based modules, often with therapist guidance via email or phone. Hundreds of randomized trials have demonstrated that ICBT is similarly effective to face-to-face psychotherapies for treating depression and anxiety [ 1 ]. There are 2 key advantages of ICBT and other digital mental health interventions (DMHIs): the ability of clients to access them privately and conveniently at practically any time and location [ 2 - 4 ] and the tendency for DMHIs to require less therapist time per client than traditional face-to-face psychotherapies [ 2 , 4 ].
ICBT is sometimes offered in a purely self-guided format (ie, without a therapist). Meta-analyses have shown that self-guided ICBT is at least slightly less effective than therapist-guided ICBT; this assertion is based on differences in effect sizes observed in separate meta-analyses of guided [ 1 ] and self-guided [ 5 , 6 ] ICBT, meta-analyses including subgroup analyses of both guided and self-guided ICBT [ 7 - 9 ], a meta-regression in which human contact predicted more favorable ICBT outcomes [ 10 ], a meta-analysis of randomized trials directly comparing guided and self-guided ICBT [ 11 ], and an individual-participant meta-analysis evaluating both guided and self-guided ICBT for depression [ 12 ]. In addition, client engagement with self-guided ICBT and other self-guided DMHIs tends to be low [ 13 - 15 ], particularly in real-world observational research, where completion rates were found in one systematic review to range from 0.5% to 28.6% [ 14 ]. However, self-guided ICBT and other self-guided DMHIs can be implemented on a large scale with minimal human or financial inputs required [ 7 , 16 ], making them cost-effective [ 17 ] and—many have argued—justifiable despite their tendency to be less effective than therapist-guided DMHIs or face-to-face psychotherapies [ 16 , 18 ].
There appears to be a growing consensus that DMHIs can be designed to be more engaging for clients [ 5 , 19 - 21 ], which may have particular implications for mitigating the problem of low engagement in self-guided DMHIs. The persuasive system design framework [ 22 ] describes 28 specific design principles for improving engagement divided into four categories: (1) primary task support principles, which facilitate completion of treatment tasks (eg, tailoring content for specific user groups and presenting complex tasks in a series of simple steps); (2) dialogue support principles, which facilitate dialogue between an intervention and its users (eg, automated praise, reminders, or virtual rewards); (3) system credibility support principles, which help ensure that users perceive interventions as credible (eg, endorsements from credible third parties and inclusion of experts in the design process); and (4) social support principles, which enable users to support each other in their use of an intervention (eg, opportunities for users to support and learn from each other). Systematic reviews and meta-analyses have shown that persuasive design characteristics can predict treatment engagement [ 20 ] and symptom change [ 5 ], but research assessing the impact of specific persuasive design principles is limited.
Online discussion forums facilitate social support principles of persuasive design. Previous research suggests that they may help support engagement and outcomes in self-guided DMHIs. For example, participants in forum-only control conditions across several studies have demonstrated promising outcomes [ 23 - 26 ], prompting the authors of one paper to conclude that they “could be regarded as an intervention” in and of themselves [ 23 ]. In total, 3 randomized trials have shown that self-guided ICBT [ 27 , 28 ] or self-guided bibliotherapy [ 3 ] accompanied by an online discussion forum exhibited equivalent outcomes to those of therapist-guided ICBT. Another randomized trial experimentally demonstrated that adding an online discussion forum to guided ICBT improved engagement [ 29 ]. Together, these studies suggest that forums could help improve engagement and outcomes in ICBT—potentially bridging the engagement and efficacy gap between guided and self-guided ICBT—but there are no previous randomized trials experimentally evaluating the impact of a forum on engagement and outcomes in self-guided ICBT.
In 2019, a clinical research unit called PSPNET was founded to develop, deliver, and conduct research on free ICBT interventions tailored specifically for Canadian first responders and other public safety personnel (PSP), who frequently experience potentially psychologically traumatic events [ 30 ], report high rates of mental health problems [ 31 ], and face unique barriers to accessing mental health care (eg, stigma within their workplaces) [ 32 , 33 ]. At the time this study was conducted, PSPNET offered 2 therapist-guided ICBT interventions to Canadian PSP—one transdiagnostic and the other posttraumatic stress disorder (PTSD) specific—both of which have shown promising outcomes with respect to symptom change, engagement, and treatment satisfaction [ 34 - 36 ]. However, at the time of this study, PSPNET was unable to offer guided ICBT services to PSP in all Canadian provinces and territories, highlighting an opportunity to develop a self-guided ICBT intervention that could be delivered with minimal resources required to PSP anywhere in Canada. No previous research has evaluated self-guided ICBT tailored specifically for PSP.
Objectives and Hypotheses
Broadly speaking, we designed this study to evaluate self-guided ICBT among Canadian PSP while addressing several questions concerning the role of online discussion forums in self-guided ICBT. Specifically, we sought to address the following four objectives:
- To evaluate transdiagnostic, self-guided ICBT tailored for PSP with respect to treatment engagement, outcomes, and satisfaction.
- To evaluate whether adding an online discussion forum to transdiagnostic, self-guided ICBT tailored for PSP would improve engagement and outcomes.
- To evaluate whether participant engagement in the online discussion forum would moderate treatment outcomes.
- To conduct a mixed methods analysis of participant feedback on the discussion forum.
We hypothesized that participants in both conditions would experience at least small to moderate reductions in symptoms of depression, anxiety, and posttraumatic stress, consistent with recent meta-analytic evidence [ 5 , 6 ]. Second, we hypothesized that participants randomly assigned to receive access to an online discussion forum would show greater engagement and more favorable treatment outcomes than those randomly assigned to receive ICBT without a discussion forum.
Study Design
We used a randomized trial design with 2 conditions: an ICBT plus peer support forum condition and an ICBT-only condition. Participants in both conditions were given free access to a self-guided ICBT program called the Self-Guided PSP Wellbeing Course . For participants in the ICBT plus peer support forum condition, but not for those in the ICBT-only condition, this ICBT course included a built-in online discussion forum. Participants were not blinded to their own condition as it is not possible to hide therapy content from those receiving therapy, but the experimental manipulation was described only in general terms (ie, without reference to forums) such that participants were blind to how the condition to which they were assigned differed from the condition to which they were not assigned. We adopted a simple randomization approach [ 37 ], which we implemented via a random number generator with a 1:1 ratio. We registered the methodological protocol for this research on ClinicalTrials.gov (ID NCT05145582) and made 2 deviations from it. First, we removed the Sheehan Disability Scale [ 38 ] from our planned outcome measures because we were unable to obtain permission to use it. Second, we ultimately carried out our primary quantitative analyses using multilevel modeling (MLM) instead of generalized estimating equations, as we had originally planned, because a paper was published during the course of this research that provided a compelling rationale and detailed recommendations for using MLM in treatment-control pretest-posttest-follow-up study designs [ 39 ]. We followed the CONSORT (Consolidated Standards of Reporting Trials) guidelines [ 40 ] in reporting the findings of this research. This research was conducted within the context of author HCM’s doctoral dissertation, and we would refer interested readers to this dissertation (expected to be publicly available in or around October 2024) for further details about this research.
This study was conducted in Canada, where publicly funded mental health services have not met public demand, leading many Canadians to access private mental health care instead [ 41 ]. Canadians have access to DMHIs through various Canadian organizations [ 42 ]. There are also thousands of mental health–related phone apps and websites available in Canada and other countries [ 43 ], but many of these services are not empirically supported. All research activities pertaining to this study were carried out at the University of Regina in Saskatchewan, Canada.
Participants, Recruitment, and Enrollment
A power analysis indicated that a minimum of 110 participants would be required to achieve adequate power to detect moderate between-group differences (see Multimedia Appendix 1 [ 3 , 5 , 8 , 27 , 34 , 44 - 47 ] for details on our sample size planning). Participants were informed about this research via paid social media advertisements (ie, Twitter [subsequently rebranded X] and Facebook), emails forwarded to PSP by leaders of PSP organizations, presentations to PSP organizations by author HCM and PSPNET’s clinicians, and word of mouth. To be eligible to take part, prospective participants were required to self-report (1) being aged ≥18 years; (2) residing in Canada; (3) working, volunteering, or having previously worked or volunteered as a PSP; (4) being able to access the internet via a computer; and (5) not experiencing significant ongoing concerns related to alcohol or drug use, psychotic symptoms, or manic symptoms.
Prospective participants accessed this study through a web page on PSPNET’s website, which provided information about the study. Upon reviewing a consent form and consenting to participate, they accessed a series of eligibility screening questionnaires through Qualtrics (Qualtrics International Inc). We contacted prospective participants by email to inform them of their eligibility, and eligible participants were asked to confirm their intent to take part in the study, after which they were randomly assigned to 1 of the 2 conditions and provided with a temporary password to access the version of the Self-Guided PSP Wellbeing Course (ie, with or without the peer support forum) to which they had been assigned. All randomization and enrollment procedures were carried out by author HCM and research assistant Julia Gregory (see the Acknowledgments section). Recruitment took place between December 6, 2021, and September 26, 2022.
Primary Outcome Measures
Patient health questionnaire–9.
The Patient Health Questionnaire–9 (PHQ-9) is a psychometrically sound, 9-item questionnaire assessing depressive symptoms [ 48 , 49 ]. Possible total scores range from 0 to 27, and a score of ≥10 suggests that a respondent’s symptoms are clinically significant [ 50 ].
Generalized Anxiety Disorder–7
The Generalized Anxiety Disorder–7 (GAD-7) is a 7-item questionnaire assessing generalized anxiety that has demonstrated strong psychometric properties [ 49 , 51 ]. Total scores can range from 0 to 21, with a score of ≥10 suggesting clinically significant symptoms [ 49 , 51 ].
PTSD Checklist for DSM-5
PTSD Checklist for DSM-5 (PCL-5) is a psychometrically sound questionnaire assessing PTSD symptoms [ 52 ]. Responses to its 20 items sum to a total score ranging from 0 to 80, and a score of ≥33 indicates that a respondent likely meets criteria for a PTSD diagnosis [ 53 , 54 ].
Secondary Outcome Measures
Brief resilience scale.
The Brief Resilience Scale (BRS) is a 6-item questionnaire measure of resilience that has shown good psychometric properties [ 55 , 56 ]. Each item has 5 response options with associated numerical values ranging from 1 to 5, and a higher mean score across items indicates greater resilience.
Flourishing Scale
The Flourishing Scale (FS) is an 8-item questionnaire assessing flourishing across various domains of life (eg, relationships, meaning and purpose, and feeling of competence). It has demonstrated good psychometric properties [ 57 , 58 ]. Total scores can range from 8 to 56, with greater scores indicating a greater degree of flourishing.
Treatment Satisfaction Questionnaire
We administered a bespoke questionnaire designed to assess treatment satisfaction and solicit feedback on the Self-Guided PSP Wellbeing Course through a mix of yes or no, Likert-scale, and open-ended text response items. For participants in the ICBT plus peer support forum condition, the Treatment Satisfaction Questionnaire included several additional items pertaining to the forum. Open-ended questions about both the course and the forum were designed to solicit both positive and constructive feedback.
Adapted Session Rating Scale
We administered a modified version of the Session Rating Scale (SRS), a 4-item questionnaire originally designed to assess client perspectives on the quality of the therapeutic alliance in face-to-face therapy [ 59 ]. It has good psychometric qualities [ 60 , 61 ]. Items are rated on a 7-point Likert scale from 0 ( absolutely disagree ) to 6 ( absolutely agree ) and assess the therapeutic bond, goal agreement, task agreement, and overall alliance quality. Following an approach taken in another study [ 62 ], we adapted the SRS to measure patient-program alliance.
Program Use Questionnaire
We administered a brief bespoke questionnaire assessing engagement with the Self-Guided PSP Wellbeing Course and, if applicable, the peer support forum. Specifically, this questionnaire was designed to assess effort put into the course; the perceived helpfulness of the course; and, if applicable, use and perceived helpfulness of the peer support forum. Program use patterns were also assessed via automatic collection of program use data (eg, number of lessons and additional resources accessed).
Health Service Use Questionnaire
We also administered a bespoke questionnaire to assess use of health care services for mental health challenges during eligibility screening and at 8 and 20 weeks after enrollment. In the interest of brevity, and because this questionnaire is peripheral to the primary objectives of this study, we do not describe the outcomes of this questionnaire in this paper.
Pre-Enrollment Measures
During eligibility screening, we administered a bespoke participant information questionnaire assessing demographic, occupational, and clinical characteristics; an ICBT feedback questionnaire assessing pre-enrollment knowledge and attitudes toward ICBT; the Credibility/Expectancy Questionnaire (CEQ) [ 63 ]; the Alcohol Use Disorders Identification Test [ 64 ]; and the Drug Use Disorders Identification Test [ 65 ].
Administration of Measures
During eligibility screening, we administered the PHQ-9, GAD-7, PCL-5, BRS, FS, participant information questionnaire, ICBT feedback questionnaire, CEQ, Alcohol Use Disorders Identification Test, and Drug Use Disorders Identification Test. At 2, 4, 6, and 8 weeks after enrollment, we administered the Program Use Questionnaire. At 8 weeks after enrollment, we also administered the PHQ-9, GAD-7, PCL-5, BRS, FS, and Treatment Satisfaction Questionnaire. At 20 weeks after enrollment, we administered the PHQ-9, GAD-7, PCL-5, BRS, and FS. Our research team encouraged participants to complete the questionnaires via emails and phone calls but did not urge participants to use the Self-Guided PSP Wellbeing Course .
Intervention
The Self-Guided PSP Wellbeing Course is an 8-week self-guided, transdiagnostic ICBT program that can be accessed through a web browser. It includes 5 core lessons, each consisting of a welcome video, a series of slides with instructive text and diagrams, an audio file covering the same clinical content as the lesson slides, illustrative case stories, frequently asked questions, downloadable homework activities called “DIY Guides,” and quotes from previous clients. These lessons included an introduction to the cognitive behavioral model and psychoeducation to help participants recognize and understand their symptoms (lesson 1); skills to help participants recognize and challenge unhelpful thoughts (lesson 2); skills for managing physiological underarousal and overarousal symptoms (lesson 3); skills for managing behavioral symptoms (lesson 4); and strategies for maintaining treatment gains, setting goals, and preventing future relapses (lesson 5). The course also included 14 additional resources covering a wide range of topics (eg, assertiveness, physical pain, and intimate relationships) and automated email reminders to encourage engagement.
The Self-Guided PSP Wellbeing Course is effectively a self-guided version of a previously developed therapist-guided ICBT course called the PSP Wellbeing Course [ 34 , 36 ]; aside from the provision of therapist guidance in the latter but not the former, the courses are practically identical. PSPNET developed the PSP Wellbeing Course by tailoring an existing ICBT program called the Wellbeing Course to meet the needs of Canadian PSP based on feedback provided by Canadian PSP in a series of interviews, focus groups, and questionnaires [ 33 , 66 ]. The original Wellbeing Course was initially developed by the eCentreClinic at Macquarie University, Australia, and has since shown excellent outcomes among the general population in Australia [ 67 ] and Canada [ 68 ].
Discussion Forum
The peer support forum was built into the Self-Guided PSP Wellbeing Course . It included 11 sections (eg, one for each of the 5 lessons, one for discussing families and relationships, and one for discussing workplace issues). It was monitored daily and moderated as required each business day by author HCM, who posed questions to spark discussion and responded to participants’ posts.
Ethical Considerations
This study was approved by the University of Regina Research Ethics Board (file 2021-130). Before taking part, all participants were provided with an informed consent form, which described the following: the objectives of the research, the research team, what participation would entail (ie, the intervention and questionnaires), possible risks and benefits of participating, project funding, considerations regarding concurrent mental health treatments, right to withdraw, limits to confidentiality, risks to privacy, precautions to improve security of participant information (both PSPNET’s precautions and precautions that participants could take), uses of participants’ data (ie, eligibility determination and research), information on accessing research results, a statement indicating that participants would not be compensated for taking part, and an invitation to contact our team with any questions or concerns. All participant data were deidentified before analysis. Due to ethical concerns related to the exclusion of individuals reporting suicidal ideation from DMHI research [ 69 ], we tried to refer prospective participants reporting suicidal ideation to more intensive services and clarified that the Self-Guided PSP Wellbeing Course is not a crisis service, but we allowed them to participate if they met the eligibility criteria.
Quantitative Data Analyses
We carried out all quantitative analyses using SPSS (version 28; IBM Corp). We did not statistically test for group differences in pre-enrollment variables as it is not meaningful to test the probability that group differences occurred by chance when it is already known—due to random assignment—that they did [ 70 ]. Instead, we inspected the magnitude of group differences and planned to conduct sensitivity analyses to assess the impact of marked differences should we observe any. We compared changes in scores on the PHQ-9, GAD-7, PCL-5, FS, and BRS across conditions using an MLM approach recommended by Sharpe and Cribbie [ 39 ]. We used an intention-to-treat approach [ 71 ] including all participants in the analyses, and we accounted for missing data using the restricted maximum likelihood estimation method, which previous research suggests is preferable to maximum likelihood estimation for MLM when random effects are included [ 72 ]. Each model was run using a random intercept and fixed effects of group, time (as a categorical variable), and the interaction between group and time. We used a variance components covariance structure [ 73 ]. We also produced a G matrix for each model consistent with the recommendations by Sharpe and Cribbie [ 39 ]. We used scatterplots and histograms to test the assumptions of linearity, homoscedasticity of residuals, and normality of residuals [ 74 ]. For each of the 5 outcome variables, we conducted one model for the entire sample and one model for the subset of participants with clinically significant scores at the pre-enrollment time point, which we defined using established cutoff scores for the PHQ-9 (≥10), GAD-7 (≥10), or PCL-5 (≥33) and scores in the lower 3 quartiles on the FS (<48) and BRS (<4.0). Therefore, we ran 10 models in total.
In each of the 10 MLM models, we conducted 5 contrasts. In total, 2 contrasts were designed to assess for interactions between group and time—that is, to identify any differences between groups with respect to changes in dependent variables over time—including one contrast for the period between the pre-enrollment time point and 8 weeks after beginning treatment and one for the period between the pre-enrollment time point and 20 weeks after beginning treatment. We collapsed the 2 groups for 3 additional contrasts to determine whether changes in questionnaire scores over time were statistically significant—one contrast for the period between the pre-enrollment time point and 8 weeks after beginning treatment, one for the period between the pre-enrollment time point and 20 weeks after beginning treatment, and one for the period between 8 weeks and 20 weeks after beginning treatment. These latter 3 contrasts were designed to assess whether participants in the Self-Guided PSP Wellbeing Course experienced significant changes in their mental health.
Finally, in each of the 10 MLM models, we investigated the effects of five covariates on changes in questionnaire scores over time: (1) the number of lessons that participants accessed, (2) the number of additional resources that participants accessed, (3) CEQ credibility scores, (4) CEQ expectancy scores, and (5) gender. These analyses are not central to the objectives of this study but may be of interest to some readers; accordingly, a rationale for the inclusion of these specific covariates, methods and results pertaining to our covariate analyses, and a brief discussion of the findings of those analyses are shown in Multimedia Appendix 2 [ 63 , 75 - 80 ].
In addition to the MLM models, we used 2-tailed independent-sample t tests and chi-square tests to assess for group differences in treatment satisfaction and program use. Upon observing possible group differences in rates of questionnaire completion, we conducted additional (non-prespecified) chi-square tests to evaluate their significance.
Qualitative Data Analyses
We conducted qualitative analyses using a content analysis approach to explore participant feedback on the peer support forum and the Self-Guided PSP Wellbeing Course in general [ 81 ]. After removing identifying information from the data, author HCM identified categories using a descriptive, inductive approach and grouped those categories into overarching themes. Given the relatively small amount of data, this was carried out using an Excel (Microsoft Corp) spreadsheet. The initial codebook was refined through meeting with author HDH and Dr Janine Beahm (see the Acknowledgments section).
It is a common practice for researchers using qualitative methods to engage in reflexivity, which is a practice of reflection on how the researchers’ positionality might affect the process or outcomes of qualitative research [ 82 ]. Being neither PSP nor ICBT clients, the authors do not identify as members of the population under study, potentially granting the authors a degree of neutrality in describing participants’ reported experiences but also potentially impeding their ability to fully understand those experiences [ 82 ]. In addition, the authors held certain attitudes and beliefs (eg, that ICBT can be helpful for many people and that forums may be able to enhance ICBT) that may have influenced the process and outcomes of this research. Nevertheless, we endeavored to minimize the risk of bias in this research by (1) including neutrally worded questions to solicit both positive and negative feedback; (2) conducting content analysis as descriptively as possible and avoiding even minor inferences and assumptions; (3) separating qualitative data from other data that could cause bias in coding (eg, demographic and clinical characteristics) before analysis; and (4) involving 3 researchers, as noted previously, in checking the accuracy of our coding.
Participants
Of the 188 prospective participants who completed the web-based screening, 153 (81.4%) were enrolled in the study and randomized, and 107 (56.9%) were included in our analyses. A flowchart displaying enrollment, program use, and questionnaire completion is shown in Figure 1 . Of note, Figure 1 shows that 36 participants in the ICBT-only condition completed symptom measures at 20 weeks after enrollment; one of these participants completed the PHQ-9 and GAD-7 but not the PCL-5. Participant characteristics are shown in Table 1 . Chi-square tests evidenced that the difference between conditions with respect to the proportion of participants who completed posttreatment questionnaires was statistically significant at 8 weeks (n=107, χ 2 1 =6.4, P =.01) but not at 20 weeks (n=107, χ 2 1 =0.5, P =.47).

| Characteristics | All participants | ICBT -only condition (n=51) | ICBT plus peer support forum condition (n=56) | |
| Women | 62 (57.9) | 28 (54.9) | 34 (60.7) | |
| Men | 45 (42.1) | 23 (45.1) | 22 (39.3) | |
| Married, common-law marriage, or living with a partner | 78 (72.9) | 38 (74.5) | 40 (71.4) | |
| Not married, in a common-law marriage, or living with a partner | 29 (27.1) | 13 (25.5) | 16 (28.6) | |
| Has ≥1 children | 70 (65.4) | 36 (70.6) | 34 (60.7) | |
| Has no children | 37 (34.6) | 15 (29.4) | 22 (39.3) | |
| British Columbia | 23 (21.5) | 13 (25.5) | 10 (17.9) | |
| Ontario | 21 (19.6) | 10 (19.6) | 11 (19.6) | |
| Alberta | 15 (14) | 9 (17.6) | 6 (10.7) | |
| New Brunswick | 12 (11.2) | 4 (7.8) | 8 (14.3) | |
| Nova Scotia | 11 (10.3) | 4 (7.8) | 7 (12.5) | |
| Prince Edward Island | 10 (9.3) | 5 (9.8) | 5 (8.9) | |
| Saskatchewan | 8 (7.5) | 4 (7.8) | 4 (7.1) | |
| Manitoba | 4 (3.7) | 0 (0) | 4 (7.1) | |
| Newfoundland and Labrador | 1 (0.9) | 1 (2) | 0 (0) | |
| Northwest Territories | 1 (0.9) | 1 (2) | 0 (0) | |
| Quebec | 1 (0.9) | 0 (0) | 1 (1.8) | |
| <100,000 residents | 72 (67.3) | 32 (62.7) | 40 (71.4) | |
| ≥100,000 residents | 35 (32.7) | 19 (37.3) | 16 (28.6) | |
| No university degree | 56 (52.3) | 26 (51) | 30 (53.6) | |
| University degree | 51 (47.7) | 25 (49) | 26 (46.4) | |
| , n (%) | ||||
| ≥10 | 75 (70.1) | 37 (72.5) | 38 (67.9) | |
| 0-9 | 32 (29.9) | 14 (27.5) | 18 (32.1) | |
| Police | 37 (34.6) | 19 (37.3) | 18 (32.1) | |
| Corrections | 23 (21.5) | 10 (19.6) | 13 (23.2) | |
| Paramedics or related emergency service | 16 (15) | 9 (17.6) | 7 (12.5) | |
| Fire | 11 (10.3) | 5 (9.8) | 6 (10.7) | |
| Communications (eg, 911 dispatch) | 7 (6.5) | 2 (3.9) | 5 (8.9) | |
| Other | 13 (12.1) | 6 (11.8) | 7 (12.5) | |
| Indigenous (ie, First Nations, Inuit, or Metis) | 7 (6.5) | 6 (11.8) | 1 (1.8) | |
| White | 96 (89.7) | 42 (82.4) | 54 (96.4) | |
| Other ethnic minority group | 3 (2.8) | 2 (3.9) | 1 (1.8) | |
| Prefer not to answer | 1 (0.9) | 1 (2) | 0 (0) | |
| 20-29 | 7 (6.5) | 4 (7.8) | 3 (5.4) | |
| 30-39 | 27 (25.2) | 16 (31.4) | 11 (19.6) | |
| 40-49 | 47 (43.9) | 22 (43.1) | 25 (44.6) | |
| 50-59 | 21 (19.6) | 9 (17.6) | 12 (21.4) | |
| 60-69 | 5 (4.7) | 0 (0) | 5 (8.9) | |
| Age (y), mean (SD) | 44.50 (9.28) | 42.97 (8.94) | 45.90 (9.45) | |
a ICBT: internet-delivered cognitive behavioral therapy.
b PSP: public safety personnel.
Changes in Questionnaire Scores
We observed a common pattern of results across all 10 MLM models: (1) all statistical assumptions were met; (2) we did not identify statistically significant effects of group or group-by-time interactions ( P ≥.17 in all cases); (3) contrasts showed no effect of group on score change at 8 or 20 weeks for any measure; (4) there was a statistically significant and favorable effect of time (ie, scores on the PHQ-9, GAD-7, and PCL-5 decreased and scores on the FS and BRS increased over time); (5) contrasts showed statistically significant improvement in scores at 8 and 20 weeks for all measures; and (6) we identified residual variance, suggesting that models were likely missing predictor variables that could have helped account for estimates of dependent variables (which was expected given that covariates were tested separately via contrasts rather than being included in MLM models).
There were also some differences across MLM models. Certain contrasts for the PHQ-9 and PCL-5 showed further improvement in symptoms from 8 to 20 weeks. Further details of MLM results are reported in Table 2 (estimated means and percentage changes) and Table 3 (contrasts). Unaltered questionnaire scores observed among respondents at each time point are shown in Multimedia Appendix 3 [ 83 - 86 ].
| Questionnaires and time points | Entire sample | Clinical subsamples | ||||||
| Both conditions | ICBT only | ICBT plus peer support forum | Both conditions | ICBT only | ICBT plus peer support forum | |||
| Pre-enrollment time point, estimated mean | 9.51 | 9.69 | 9.34 | 14.21 | 14.35 | 14.07 | ||
| 8 weeks, estimated mean (% change from pre-enrollment time point) | 7.26 (–23.7) | 6.96 (–28.2) | 7.53 (–19.4) | 9.58 (–32.6) | 9.86 (–31.3) | 9.31 (–33.9) | ||
| 20 weeks, estimated mean (% change from pre-enrollment time point) | 6.01 (–36.7) | 5.94 (–38.7) | 6.08 (–34.9) | 7.94 (–44.1) | 7.96 (–44.5) | 7.93 (–43.6) | ||
| Pre-enrollment time point, estimated mean | 8.10 | 8.34 | 7.88 | 13.79 | 14.47 | 13.15 | ||
| 8 weeks, estimated mean (% change from pre-enrollment time point) | 6.19 (–23.5) | 5.92 (–29.0) | 6.44 (–18.3) | 8.71 (–36.9) | 9.08 (–37.3) | 8.35 (–36.5) | ||
| 20 weeks, estimated mean (% change from pre-enrollment time point) | 5.31 (–34.5) | 5.47 (–34.4) | 5.16 (–34.4) | 8.25 (–40.2) | 8.46 (–41.6) | 8.05 (–38.8) | ||
| Pre-enrollment time point, estimated mean | 26.86 | 24.73 | 28.80 | 46.64 | 47.47 | 46.08 | ||
| 8 weeks, estimated mean (% change from pre-enrollment time point) | 18.71 (–30.3) | 17.61 (–28.8) | 19.71 (–31.6) | 28.68 (–38.5) | 33.84 (–28.7) | 25.17 (–45.4) | ||
| 20 weeks, estimated mean (% change from pre-enrollment time point) | 16.49 (–38.6) | 15.52 (–37.2) | 17.37 (–39.6) | 23.67 (–49.3) | 24.78 (–47.8) | 22.91 (–50.3) | ||
| Pre-enrollment time point, estimated mean | 40.83 | 40.92 | 40.75 | 37.63 | 37.16 | 38.02 | ||
| 8 weeks, estimated mean (% change from pre-enrollment time point) | 42.22 (3.4) | 41.86 (2.3) | 42.55 (4.4) | 39.78 (5.7) | 38.49 (3.6) | 40.86 (7.5) | ||
| 20 weeks, estimated mean (% change from pre-enrollment time point) | 43.35 (6.2) | 43.28 (5.8) | 43.41 (6.5) | 41.11 (9.3) | 40.34 (8.6) | 41.76 (9.8) | ||
| Pre-enrollment time point, estimated mean | 3.28 | 3.33 | 3.24 | 2.92 | 2.96 | 2.88 | ||
| 8 weeks, estimated mean (% change from pre-enrollment time point) | 3.51 (6.8) | 3.47 (4.0) | 3.54 (9.3) | 3.29 (12.9) | 3.32 (12.3) | 3.27 (13.5) | ||
| 20 weeks, estimated mean (% change from pre-enrollment time point) | 3.48 (5.9) | 3.55 (6.5) | 3.41 (5.3) | 3.26 (11.8) | 3.33 (12.5) | 3.20 (11.0) | ||
b PHQ-9: Patient Health Questionnaire–9.
c Entire sample—both conditions: n=107, ICBT-only condition: n=51, and ICBT plus peer support forum: n=56; clinical subsamples—both conditions: n=53, ICBT-only condition: n=26, and ICBT plus peer support forum: n=27.
d GAD-7: Generalized Anxiety Disorder–7.
e Entire sample—both conditions: n=107, ICBT-only condition: n=51, and ICBT plus peer support forum: n=56; clinical subsamples—both conditions: n=39, ICBT-only condition: n=19, and ICBT plus peer support forum: n=20.
f PCL-5: PTSD Checklist for DSM-5.
g Entire sample—both conditions: n=107, ICBT-only condition: n=51, and ICBT plus peer support forum: n=56; clinical subsamples—both conditions: n=42, ICBT-only condition: n=17, and ICBT plus peer support forum: n=25.
h FS: Flourishing Scale.
i Entire sample—both conditions: n=107, ICBT-only condition: n=51, and ICBT plus peer support forum: n=56; clinical subsamples—both conditions: n=81, ICBT-only condition: n=37, and ICBT plus peer support forum: n=44.
j BRS: Brief Resilience Scale.
k Entire sample—both conditions: n=107, ICBT-only condition: n=51, and ICBT plus peer support forum: n=56; clinical subsamples—both conditions: n=78, ICBT-only condition: n=37, and ICBT plus peer support forum: n=41.
| Variables | Entire sample | Clinical subsamples | ||||||
| test ( ) | value | Cohen | test ( ) | value | Cohen | |||
| Pre-enrollment time point to 8 weeks after beginning treatment | –4.44 (158.78) | <.001 | –0.70 | –6.80 (76.52) | <.001 | –1.55 | ||
| Pre-enrollment time point to 20 weeks after beginning treatment | –6.55 (160.38) | <.001 | –1.03 | –8.39 (78.87) | <.001 | –1.89 | ||
| 8-20 weeks after beginning treatment | –2.20 (156.57) | .03 | –0.35 | –2.11 (77.13) | .04 | –0.48 | ||
| Pre-enrollment time point to 8 weeks after beginning treatment | –4.18 (158.10) | <.001 | –0.66 | –6.67 (54.53) | <.001 | –1.81 | ||
| Pre-enrollment time point to 20 weeks after beginning treatment | –5.77 (159.88) | <.001 | –0.91 | –6.56 (56.78) | <.001 | –1.74 | ||
| 8-20 weeks after beginning treatment | –1.71 (155.65) | .09 | –0.27 | –0.52 (54.08) | .61 | –0.14 | ||
| Pre-enrollment time point to 8 weeks after beginning treatment | –5.67 (151.91) | <.001 | –0.92 | –8.39 (57.98) | <.001 | –2.20 | ||
| Pre-enrollment time point to 20 weeks after beginning treatment | –6.88 (152.70) | <.001 | –1.11 | –10.58 (58.75) | <.001 | –2.76 | ||
| 8-20 weeks after beginning treatment | –1.42 (149.50) | .16 | –0.23 | –2.51 (56.12) | .02 | –0.67 | ||
| Pre-enrollment time point to 8 weeks after beginning treatment | 2.10 (159.08) | .04 | 0.33 | 2.61 (118.39) | .01 | 0.48 | ||
| Pre-enrollment time point to 20 weeks after beginning treatment | 3.67 (159.97) | <.001 | 0.58 | 4.11 (119.64) | <.001 | 0.75 | ||
| 8-20 weeks after beginning treatment | 1.60 (157.12) | .11 | 0.26 | 1.55 (117.03) | .12 | 0.29 | ||
| Pre-enrollment time point to 8 weeks after beginning treatment | 3.00 (160.80) | .003 | 0.47 | 4.94 (115.02) | <.001 | 0.92 | ||
| Pre-enrollment time point to 20 weeks after beginning treatment | 2.55 (162.06) | .01 | 0.40 | 4.23 (116.22) | <.001 | 0.78 | ||
| 8-20 weeks after beginning treatment | –0.29 (158.65) | .77 | –0.05 | –0.39 (112.77) | .70 | –0.07 | ||
a PHQ-9: Patient Health Questionnaire–9.
b Entire sample: n=107; clinical subsamples: n=53.
c GAD-7: Generalized Anxiety Disorder–7.
d Entire sample: n=107; clinical subsamples: n=39.
e PCL-5: PTSD Checklist for DSM-5.
f Entire sample: n=107; clinical subsamples: n=42.
g FS: Flourishing Scale.
h Entire sample: n=107; clinical subsamples: n=81.
i BRS: Brief Resilience Scale.
j Entire sample: n=107; clinical subsamples: n=78.
Program Use
There was no statistically significant difference between groups with respect to the number of lessons participants accessed by 8 weeks (t 105 =–0.28; P= .78; Cohen d =–0.05) or 20 weeks (t 105 =0.82; P= .42; Cohen d =0.16). Collapsing across groups, a sizeable minority of participants accessed all 5 lessons of the Self-Guided PSP Wellbeing Course by 8 weeks (30/107, 28%) or 20 weeks (46/107, 43%). Nearly half (48/107, 44.9%) accessed 4 of 5 lessons by 8 weeks, whereas more than half (59/107, 55.1%) accessed 4 of 5 lessons by 20 weeks. Participants accessed an average of 3.33 (SD 5.00) additional resources. Responses to the Program Use Questionnaire collapsed across groups and averaged across time points (ie, 2, 4, 6, and 8 weeks) showed that participants most commonly reported putting “some effort” into the course (39%), followed by “a little effort” (30.9%), “no effort” (17.5%), and “a lot of effort” (12.5%), with no participants reporting “a great deal of effort” at any time point.
Treatment Satisfaction
The Treatment Satisfaction Questionnaire and SRS were completed by 68% (38/56) of the participants in the ICBT plus peer support forum condition and 88% (45/51) of the participants in the ICBT-only condition. Two-tailed independent-sample t tests and chi-square tests showed no statistically significant differences between groups with respect to any treatment satisfaction variables ( P ≥.47 in all cases). Accordingly, we present the results collapsed across groups in Table 4 .
We qualitatively analyzed responses to open-ended questions from 61.7% (66/107) of the participants, which we organized into 3 main themes: positive feedback, negative or constructive feedback, and comments about personal circumstances or preferences that do not reflect the perceived helpfulness of the Self-Guided PSP Wellbeing Course . The results are shown in Table 5 .
| Variables | Values | |
| Yes | 79 (96) | |
| No | 3 (4) | |
| Yes | 78 (94) | |
| No | 5 (6) | |
| Very dissatisfied (0), n (%) | 0 (0) | |
| Dissatisfied (1), n (%) | 0 (0) | |
| Neutral (2), n (%) | 31 (37) | |
| Satisfied (3), n (%) | 45 (54) | |
| Very satisfied (4), n (%) | 7 (8) | |
| Values, mean (SD) | 2.71 (0.62) | |
| Greatly reduced (0), n (%) | 3 (4) | |
| Reduced (1), n (%) | 3 (4) | |
| No change (2), n (%) | 21 (26) | |
| Increased (3), n (%) | 49 (60) | |
| Greatly increased (4), n (%) | 6 (7) | |
| Values, mean (SD) | 2.63 (0.82) | |
| Strongly disagree | 2 (2) | |
| Disagree | 14 (17) | |
| Neutral | 32 (39) | |
| Agree | 27 (33) | |
| Strongly agree | 7 (9) | |
| Strongly disagree | 3 (4) | |
| Disagree | 19 (23) | |
| Neutral | 32 (39) | |
| Agree | 20 (24) | |
| Strongly agree | 8 (10) | |
| , mean (SD) | ||
| Bond (n=83) | 4.55 (1.13) | |
| Goals (n=82) | 4.67 (1.01) | |
| Tasks (n=82) | 4.37 (1.50) | |
| Overall (n=82) | 4.55 (1.29) | |
a SRS: Session Rating Scale.
| Theme, subtheme, and category | Example quote | Frequency, n (%) | ||
| Positive feedback on stories or case examples | “I liked the stories cause it helped me relate and see other people are having these experiences.” [Participant 1801] | 18 (27) | ||
| Positive feedback on DIY guides | “DIY Guides are very informative and easy to understand.” [Participant 1256] | 17 (26) | ||
| Positive feedback on additional resources | “I liked the resource library to be accessed for follow up and reminders.” [Participant 1757] | 13 (20) | ||
| Positive feedback on course content (eg, thorough, understandable, and relatable) | “It goes into explanations that in person therapy doesn’t seem to have time for, or, that in person therapists don’t think to cover.” [Participant 1392] | 8 (12) | ||
| Positive feedback on lessons | “Lessons were straight forward and easy to comprehend.” [Participant 1583] | 5 (8) | ||
| Positive feedback on tools and skills taught in the course | “Gave me a framework to understand what has been affecting me and how to work on it productively. I have taken my time, more than intended by the course I think, to practice the skills.” [Participant 1175] | 6 (9) | ||
| Course acted as a helpful reminder of previously learned skills and information | “Good refresher and reminder of important concepts.” [Participant 1342] | 4 (6) | ||
| Liked that the course was self-guided, self-paced, or accessible at any time and location | “[Liked] being able to work on the course on my own timeline, when I was in the right headspace. It didn’t feel forced.” [Participant 1091] | 12 (18) | ||
| Liked the format or structure of the course or the presentation or delivery of information | “I liked how the course was structured.” [Participant 1648] | 10 (15) | ||
| Liked being able to download or print course materials or review them again in the future | “It is nice to have the resources to go back to in the future.” [Participant 1648] | 5 (8) | ||
| Liked the reminder emails | “[Liked] reminders to keep at it.” [Participant 1225] | 3 (5) | ||
| General statement of liking the course | “[Liked] honestly, all of it.” [Participant 1092] | 2 (3) | ||
| No positive feedback provided | — | 2 (3) | ||
| Disliked the stories, did not find them helpful, or provided feedback on them | “I didn’t find the stories particularly helpful.” [Participant 1154] | 5 (8) | ||
| Course was too basic or recommendation for a second course with more tools | “I thought it would be longer and more in depth.” [Participant 1742] | 3 (5) | ||
| Some content seemed redundant or unnecessary | “I found the lessons and DIY guides a bit repetitive (they covered a lot of the same material).” [Participant 1173] | 2 (3) | ||
| Other suggestions for improving clinical content | “I was approaching this as a preventative course as opposed to a treatment course so I found that the examples were not something I identified with. It would be wonderful if there was a separate course for individuals looking to build skills to help prevent a slide into negative mental health.” [Participant 1258] | 3 (5) | ||
| Difficulty or dislike concerning the current use of timelines and reminders to motivate completion | “I needed more time and felt somewhat anxious when the reminders were coming about a new section and I was behind.” [Participant 1503] | 7 (11) | ||
| Would prefer if the course included therapist support | “I wished I also had the therapist to help keep me on track and discuss some of my thoughts and feelings that came up while taking the course.” [Participant 1801] | 5 (8) | ||
| Disliked amount of reading or suggested more video content | “[Disliked] a lot of reading. Hard to stay focused.” [Participant 1546] | 3 (5) | ||
| Course was not mobile friendly | “The slides were difficult to read in a phone. Sitting at a computer isn’t always an option for privacy.” [Participant 1181] | 1 (2) | ||
| No dislikes identified or constructive feedback provided | “There is nothing I didn’t like.” [Participant 1242] | 29 (44) | ||
| Limited time, energy, or capacity to work on the course or unexpected life circumstances posing a barrier to progression in the course | “Nothing you can do but life threw me a curve the past couple weeks, very sick kitten so that was my immediate concern and this fell to the wayside.” [Participant 1816] | 6 (9) | ||
| Hard time with web-based courses in general, preference for in-person courses or would benefit more from in-person courses | “I would benefit more from in-person treatment, but am reluctant to participate.” [Participant 1225] | 3 (5) | ||
a DIY: do-it-yourself.
b Not applicable.
Online Discussion Forum Use and Satisfaction
Only 9% (5/56) of the participants in the ICBT plus peer support forum condition posted in the forum, creating 9 posts in total. The moderator (author HCM) created an additional 16 posts in an effort to spark discussion. The Treatment Satisfaction Questionnaire was completed by 38 participants in the ICBT plus peer support forum condition, 14 (37%) of whom reported that they did not use the forum. Of the remaining 24 participants, 1 (4%) reported feeling “very satisfied” with the forum overall, 11 (46%) reported feeling “satisfied,” 8 (33%) reported feeling “neutral,” 4 (17%) reported feeling “dissatisfied,” and none reported feeling “very dissatisfied.” Among the 38 participants in the ICBT plus peer support forum who completed the Treatment Satisfaction Questionnaire were 15 (39%) who reported reading between “a few” posts and “all or nearly all” posts, including participants who reported that reading others’ posts was “highly beneficial” (n=2, 13%), “beneficial” (n=3, 20%), “somewhat beneficial” (n=6, 40%), and “not beneficial at all” (n=4, 27%).
We received meaningful, analyzable feedback on the peer support forum from 52% (29/56) of the participants, identifying 17 categories of feedback, which we grouped into 3 general themes: positive feedback, constructive or negative feedback, and other personal reactions to the forum. The results of this content analysis are shown in Table 6 .
| Theme, subtheme, and category | Example quote | Frequency, n (%) | |||
| Liked that the forum was supportive, open, or free of judgment | “Judgement-free, supportive space to use when/if/how helpful.” [Participant 1217] | 2 (7) | |||
| Liked reading others’ comments or seeing a variety of perspectives | “[Liked] variety of viewpoints.” [Participant 1217] | 2 (7) | |||
| Liked not feeling alone | “It’s nice to know you’re not alone.” [Participant 1816] | 2 (7) | |||
| Liked that the forum was an option | “[Liked] that it was an option.” [Participant 1978] | 1 (3) | |||
| Did not like anything about the forum | “I did not [like the forum]. Possibly I didn’t connect properly?” [Participant 1721] | 2 (7) | |||
| Dislike, discomfort, or difficulty opening up to or being vulnerable with others | “[Did not post because] police culture does not encourage sharing or vulnerability with mental health. Peer forums are not a tool we are comfortable with. Especially with the association of privacy and information release in our jobs.” [Participant 1181] | 8 (28) | |||
| Disliked the low level of forum activity or did not post due to low activity level | “[Disliked that] it was not an active forum and often nothing had been posted.” [Participant 1978] | 7 (24) | |||
| Unaware of forum or comment that more prompts would result in greater forum use | “[Did not post because the forum] wasn’t emphasized enough as an available tool or resource during the course. I also did not know it was available to me.” [Participant 1095] | 4 (14) | |||
| Would prefer a scheduled live chat to asynchronous posts | “Wished it was more of a real time chat.” [Participant 1801] | 2 (7) | |||
| General statement of dislike for or disinterest in the forum | “I did not like it.” [Participant 1130] | 2 (7) | |||
| Too much involvement from the moderator | “[Disliked that the forum was]...very monitored?” [Participant 1721] | 1 (3) | |||
| Disliked nothing about the forum | “[Disliked] nothing.” [Participant 1241] | 1 (3) | |||
| Did not post because of other demands or not enough time | “[Did not post because] work and life demands paused my participation in the program.” [Participant 1584] | 4 (14) | |||
| Participant did not feel that they had anything of value to add to forum | [Did not post because] “I felt I didn’t have anything to add of value.” [Participant 1816] | 3 (10) | |||
| Comment on how it felt to post on the forum | “[It felt] very difficult, vulnerable to do, felt unburdened/heard after posting.” [Participant 1217] | 2 (7) | |||
| Misconception that participants cannot respond to each other | “[Disliked that the forum] seemed like a question and answer type without being able to respond to each other.” [Participant 1721] | 1 (3) | |||
| Did not feel a need to post because other aspects of the course were sufficient | “[Did not post because] I am still stuck on capturing my thoughts and found that the FAQ suffices.” [Participant 1168] | 1 (3) | |||
Principal Findings
ICBT is an effective mental health treatment [ 1 ], but clients demonstrate somewhat less favorable clinical outcomes [ 5 - 12 ] and engagement [ 13 - 15 ] when it is offered in a purely self-guided format. Persuasive design principles represent a possible means of improving engagement and outcomes in DMHIs [ 5 , 20 ], and preliminary evidence supports the use of social support principles of persuasive design implemented via online discussion forums [ 3 , 23 - 29 ], but there is a dearth of experimental research directly evaluating the impact of forums in ICBT or other DMHIs. Research has also shown that Canadian PSP benefit considerably from tailored, guided ICBT [ 34 - 36 ], but previous research has not evaluated self-guided ICBT among Canadian PSP. We conducted a randomized trial to assess the impact of adding an online discussion forum to self-guided ICBT and evaluate outcomes of tailored, self-guided ICBT among Canadian PSP.
Participants showed large improvements in symptoms of depression, anxiety, and posttraumatic stress, which supported and surpassed our hypothesis of at least small to moderate reductions in symptoms. Most meta-analyses of self-guided DMHIs have not reported pretest-posttest effect sizes, but our results for changes in depression over time compare favorably to the pretest-posttest effect size of d =0.78 reported in one meta-analysis of self-guided DMHIs for depression [ 87 ]. Changes in flourishing and resilience were more modest, potentially because the Self-Guided PSP Wellbeing Course was designed to reduce symptoms of mental disorders but not explicitly designed to improve flourishing or resilience. Nevertheless, the finding of improvements in flourishing and resilience makes an important contribution to the research literature as research on the effects of ICBT on these constructs is scarce.
The symptom change and treatment satisfaction demonstrated in this study were roughly comparable to those observed in research on the guided version of the PSP Wellbeing Course , likely because the 2 courses included practically identical treatment materials. However, engagement with the guided version was markedly better, with only 5.7% of enrolled PSP failing to access or withdrawing from the intervention (compared to 46/153, 30% in this study), 76.1% of participants accessing at least 4 of the 5 lessons of the course within 8 weeks (compared to 48/107, 44.9% in this study), and 57.3% completing the course within 8 weeks (compared to 30/107, 28% in this study) [ 36 ]. These findings suggest that therapists play a pivotal role in both initiating and sustaining Canadian PSP’s engagement in ICBT. This conclusion aligns with those of previous research showing that PSP very frequently cite therapist guidance as a liked aspect of ICBT [ 88 ] and with a broader research literature showing that engagement tends to be lower in self-guided than in guided DMHIs [ 13 - 15 ]. Nevertheless, engagement with the Self-Guided PSP Wellbeing Course in this study appears to compare favorably to that of other research on self-guided DMHIs. In a systematic review, Kelders et al [ 20 ] found, on average, a 54.2% rate of “intended use” (ie, some engagement but not necessarily completion) of internet interventions for mental health, including both guided and self-guided interventions. Another systematic review found real-world completion rates ranging from 0.5% to 28.6% for self-guided DMHIs [ 14 ]. Engagement in the present study may have been enhanced by the structure provided by the randomized trial design as this kind of study design was found to predict greater engagement in the review by Kelders et al [ 20 ]. Interestingly, only a minority of participants in the present study indicated that they would have preferred to receive therapist guidance via email or phone while taking the course.
The results failed to support our hypothesis that participants assigned to the ICBT plus peer support forum condition would demonstrate greater engagement and treatment outcomes. Given participants’ limited engagement with the peer support forum, this was unsurprising. The proportion of participants who posted in the forum (5/56, 9%) was far lower than proportions of 53% [ 26 ] and 50.6% [ 28 ] reported in previous studies of forums in DMHIs. Similarly, the mean number of posts per participant (0.16) was far lower than the means of 13.1 [ 89 ], 4.5 [ 89 ], 2.2 [ 29 ], and 1.86 [ 28 ] reported in previous studies. We are aware of only one previous study in which a lower proportion (ie, 7%) of participants posted in a forum accompanying a DMHI [ 90 ]. Despite low engagement with the peer support forum, some participants reported feeling satisfied with it and indicated that reading posts was beneficial, with qualitative feedback suggesting that some participants felt that the forum was supportive and helped them feel that they were not alone.
There are several possible reasons for the low engagement with the forum in this study. First, qualitative feedback suggested that many participants felt uncomfortable opening up to others and showing vulnerability, with one participant explicitly attributing this to “police culture,” suggesting that forums may be a poor fit for many PSP. Second, the fact that the Self-Guided PSP Wellbeing Course was transdiagnostic, with different participants experiencing different symptoms, may have led participants to feel that they did not have much in common with other forum users and, therefore, may also have detracted from their comfort in sharing their experiences. Third, forums may be particularly helpful as an adjunct to treatment for certain conditions; indeed, much of the past research supporting the use of forums in ICBT has been conducted within the context of ICBT for social anxiety [ 3 , 23 , 27 , 28 ]. Fourth, the PSP who self-selected into this study may have been particularly interested in independently accessing a self-guided treatment, whereas PSP who were interested in sharing their experiences with others may have opted for other mental health care options (including PSPNET’s therapist-guided ICBT for PSP in provinces where it was available). Finally, there was likely room for improvement with respect to the structure and implementation of the forum and our efforts to encourage participants to use it.
There was only one statistically significant difference observed between conditions: a greater proportion of participants completed questionnaires at 8 weeks in the ICBT-only condition. It remains unclear why this occurred. It could be a spurious finding. It could also be that participants in the ICBT plus peer support forum condition inferred from the minimal forum engagement that engagement with the study as a whole was low and were less likely to complete questionnaires due to the phenomenon of social normative influence [ 91 ].
Strengths, Limitations, and Future Research
This study benefitted from a mixed methods approach, allowing for both a quantitative evaluation of treatment outcomes and a qualitative exploration of participants’ experiences. Another strength of this study was its ecological validity as we evaluated the Self-Guided PSP Wellbeing Course and the peer support forum under the conditions in which they were designed to be implemented. This study also had important limitations. We did not include a control condition with which to compare outcomes of the course, and our inclusion of multiple outcome measures in separate analyses increased our familywise error rate. We expect that every discussion forum is unique and its social dynamics are unpredictable; therefore, a key limitation of this study is that it is, in a sense, a case study of a single forum with results that may not generalize well to other forums. This study was also sufficiently powered to detect only moderate differences between conditions, and due to an unexpectedly high rate of withdrawal from the study or failure to begin the intervention after we had ceased recruitment, we ultimately included 3 fewer participants in our analyses than originally planned. Finally, because we did not exclude participants with minimal or mild pre-enrollment symptoms from this research, floor effects are likely present in quantitative analyses conducted among our entire sample.
Future research can expand on this study and address the limitations noted previously in several ways. Although the peer support forum in this study had no demonstrable effect on treatment outcomes, previous research has shown excellent outcomes for online discussion forums [ 3 , 23 - 29 ], highlighting a need for further experimental research to evaluate the impact of forums on treatment outcomes in self-guided DMHIs. We are aware of a large factorial randomized trial assessing, among other things, the impact of an online discussion forum on treatment outcomes in ICBT, but the results of this trial are not yet available [ 92 ]. It would also be helpful for future research to identify common characteristics of forums that function well and those that do not, further explore DMHI users’ perspectives on forums, and identify strategies for improving engagement with forums drawing on the persuasive system design framework and other work. With respect to outcomes of self-guided ICBT tailored for PSP, future research could compare treatment outcomes against a control condition, assess longer-term outcomes, and assess additional outcomes that we did not assess.
Conclusions
ICBT has shown excellent outcomes for treating a range of psychological concerns among PSP [ 34 - 36 ] and the general population [ 1 ]. Self-guided ICBT is more scalable but shows poorer engagement and outcomes than therapist-guided ICBT [ 5 - 15 ]. There is emerging evidence suggesting that persuasive design may help improve engagement and outcomes in ICBT [ 5 , 19 - 21 ], but further research is needed. We conducted a randomized trial, finding that transdiagnostic self-guided ICBT tailored specifically for PSP showed good outcomes, but PSP randomly assigned to receive access to a built-in online discussion forum showed limited engagement with it and no evidence of benefitting from it. Our findings support the continued implementation of self-guided ICBT. Our findings contrast with those of previous research on discussion forums in DMHIs, which have generally shown promising engagement and outcomes [ 3 , 23 - 29 , 89 ], highlighting a need for more research to clarify the circumstances under which forums may help improve engagement and outcomes in DMHIs. More broadly, as DMHIs become increasingly popular, there is a great need for more research identifying possible strategies to make them more engaging and effective, including—but not limited to—further research evaluating the impact of specific persuasive design principles.
Acknowledgments
The authors would like to thank Dr Janine Beahm for her feedback on their qualitative analyses and Dr Donald Sharpe for his recommendations regarding their quantitative analyses. The authors would like to thank all past and current members of the PSPNET team and all public safety personnel who participated in this research for making it possible. Particular thanks are due to Julia Gregory for her work as a research assistant in carrying out this study. The authors would like to thank Drs Nick Titov and Blake Dear for sharing treatment materials used to develop the Self-Guided PSP Wellbeing Course . This research was supported by funding from the Canadian Institutes of Health Research and Public Safety Canada; neither funder had any involvement in carrying out this research. Finally, acknowledgments are due to the Public Safety Canada Steering Committee, the Canadian Institute for Public Safety Research and Treatment, the Online Therapy Unit, and Information Services at the University of Regina for supporting this research.
Data Availability
The data sets generated and analyzed during this study are not publicly available due our commitment to our participants to securely store their data and refrain from sharing them with anyone outside of our research team.
Conflicts of Interest
None declared.
Sample size planning.
Covariate analyses.
Observed descriptive statistics on questionnaire scores.
- Andersson G, Carlbring P, Titov N, Lindefors N. Internet interventions for adults with anxiety and mood disorders: a narrative umbrella review of recent meta-analyses. Can J Psychiatry. Jul 16, 2019;64(7):465-470. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Christensen H, Hickie IB. E-mental health: a new era in delivery of mental health services. Med J Aust. Jun 07, 2010;192(S11):S2-S3. [ CrossRef ] [ Medline ]
- Furmark T, Carlbring P, Hedman E, Sonnenstein A, Clevberger P, Bohman B, et al. Guided and unguided self-help for social anxiety disorder: randomised controlled trial. Br J Psychiatry. Nov 2009;195(5):440-447. [ CrossRef ] [ Medline ]
- Richards D, Enrique A, Palacios J, Duffy D. Internet-delivered cognitive behaviour therapy. In: Şenormancı Ö, Şenormancı G, editors. Cognitive Behavioral Therapy and Clinical Applications. London, UK. IntechOpen; 2018.
- McCall HC, Hadjistavropoulos HD, Sundström CR. Exploring the role of persuasive design in unguided internet-delivered cognitive behavioral therapy for depression and anxiety among adults: systematic review, meta-analysis, and meta-regression. J Med Internet Res. Apr 29, 2021;23(4):e26939. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Karyotaki E, Riper H, Twisk J, Hoogendoorn A, Kleiboer A, Mira A, et al. Efficacy of self-guided internet-based cognitive behavioral therapy in the treatment of depressive symptoms: a meta-analysis of individual participant data. JAMA Psychiatry. Apr 01, 2017;74(4):351-359. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Andersson G, Cuijpers P. Internet-based and other computerized psychological treatments for adult depression: a meta-analysis. Cogn Behav Ther. 2009;38(4):196-205. [ CrossRef ] [ Medline ]
- Wright JH, Owen JJ, Richards D, Eells TD, Richardson T, Brown GK, et al. Computer-assisted cognitive-behavior therapy for depression: a systematic review and meta-analysis. J Clin Psychiatry. Mar 19, 2019;80(2):18r12188. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Spek V, Cuijpers P, Nyklícek I, Riper H, Keyzer J, Pop V. Internet-based cognitive behaviour therapy for symptoms of depression and anxiety: a meta-analysis. Psychol Med. Mar 2007;37(3):319-328. [ CrossRef ] [ Medline ]
- Krieger T, Bur OT, Weber L, Wolf M, Berger T, Watzke B, et al. Human contact in internet-based interventions for depression: a pre-registered replication and meta-analysis of randomized trials. Internet Interv. Apr 2023;32:100617. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Baumeister H, Reichler L, Munzinger M, Lin J. The impact of guidance on internet-based mental health interventions — a systematic review. Internet Interv. Oct 2014;1(4):205-215. [ CrossRef ]
- Karyotaki E, Efthimiou O, Miguel C, Bermpohl FM, Furukawa TA, Cuijpers P, Individual Patient Data Meta-Analyses for Depression (IPDMA-DE) Collaboration, et al. Internet-based cognitive behavioral therapy for depression: a systematic review and individual patient data network meta-analysis. JAMA Psychiatry. Apr 01, 2021;78(4):361-371. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Andersson G, Carlbring P, Berger T, Almlöv J, Cuijpers P. What makes internet therapy work? Cogn Behav Ther. 2009;38 Suppl 1:55-60. [ CrossRef ] [ Medline ]
- Fleming TM, Bavin L, Lucassen M, Stasiak K, Hopkins S, Merry S. Beyond the trial: systematic review of real-world uptake and engagement with digital self-help interventions for depression, low mood, or anxiety. J Med Internet Res. Jun 06, 2018;20(6):e199. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Gan DZ, McGillivray L, Larsen ME, Christensen H, Torok M. Technology-supported strategies for promoting user engagement with digital mental health interventions: a systematic review. Digit Health. Jun 01, 2022;8:205520762210982. [ CrossRef ]
- Andersson G, Titov N. Advantages and limitations of internet-based interventions for common mental disorders. World Psychiatry. Feb 2014;13(1):4-11. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Gerhards SA, de Graaf LE, Jacobs LE, Severens JL, Huibers MJ, Arntz A, et al. Economic evaluation of online computerised cognitive-behavioural therapy without support for depression in primary care: randomised trial. Br J Psychiatry. Apr 2010;196(4):310-318. [ CrossRef ] [ Medline ]
- Lorenzo-Luaces L, Johns E, Keefe JR. The generalizability of randomized controlled trials of self-guided internet-based cognitive behavioral therapy for depressive symptoms: systematic review and meta-regression analysis. J Med Internet Res. Nov 09, 2018;20(11):e10113. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Fleming TM, de Beurs D, Khazaal Y, Gaggioli A, Riva G, Botella C, et al. Maximizing the impact of e-therapy and serious gaming: time for a paradigm shift. Front Psychiatry. 2016;7:65. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Kelders SM, Kok RN, Ossebaard HC, van Gemert-Pijnen JE. Persuasive system design does matter: a systematic review of adherence to web-based interventions. J Med Internet Res. Nov 14, 2012;14(6):e152. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Ludden GD, van Rompay TJ, Kelders SM, van Gemert-Pijnen JE. How to increase reach and adherence of web-based interventions: a design research viewpoint. J Med Internet Res. Jul 10, 2015;17(7):e172. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Oinas-Kukkonen H, Harjumaa M. Persuasive systems design: key issues, process model, and system features. Commun Assoc Inf Syst. 2009;24:485-501. [ CrossRef ]
- Andersson G, Carlbring P, Holmström A, Sparthan E, Furmark T, Nilsson-Ihrfelt E, et al. Internet-based self-help with therapist feedback and in vivo group exposure for social phobia: a randomized controlled trial. J Consult Clin Psychol. Aug 2006;74(4):677-686. [ CrossRef ] [ Medline ]
- Boettcher J, Aström V, Påhlsson D, Schenström O, Andersson G, Carlbring P. Internet-based mindfulness treatment for anxiety disorders: a randomized controlled trial. Behav Ther. Mar 2014;45(2):241-253. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Johansson P, Westas M, Andersson G, Alehagen U, Broström A, Jaarsma T, et al. An internet-based cognitive behavioral therapy program adapted to patients with cardiovascular disease and depression: randomized controlled trial. JMIR Ment Health. Oct 03, 2019;6(10):e14648. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Hesser H, Gustafsson T, Lundén C, Henrikson O, Fattahi K, Johnsson E, et al. A randomized controlled trial of internet-delivered cognitive behavior therapy and acceptance and commitment therapy in the treatment of tinnitus. J Consult Clin Psychol. Aug 2012;80(4):649-661. [ CrossRef ] [ Medline ]
- Titov N, Andrews G, Schwencke G, Solley K, Johnston L, Robinson E. An RCT comparing effect of two types of support on severity of symptoms for people completing internet-based cognitive behaviour therapy for social phobia. Aust N Z J Psychiatry. Jan 01, 2009;43(10):920-926. [ CrossRef ]
- Berger T, Caspar F, Richardson R, Kneubühler B, Sutter D, Andersson G. Internet-based treatment of social phobia: a randomized controlled trial comparing unguided with two types of guided self-help. Behav Res Ther. Mar 2011;49(3):158-169. [ CrossRef ] [ Medline ]
- Carolan S, Harris PR, Greenwood K, Cavanagh K. Increasing engagement with an occupational digital stress management program through the use of an online facilitated discussion group: results of a pilot randomised controlled trial. Internet Interv. Dec 2017;10:1-11. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Carleton RN, Afifi TO, Taillieu T, Turner S, Krakauer R, Anderson GS, et al. Exposures to potentially traumatic events among public safety personnel in Canada. Can J Behav Sci. Jan 2019;51(1):37-52. [ CrossRef ]
- Carleton RN, Afifi TO, Turner S, Taillieu T, Duranceau S, LeBouthillier DM, et al. Mental disorder symptoms among public safety personnel in Canada. Can J Psychiatry. Jan 28, 2018;63(1):54-64. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Ricciardelli R, Carleton RN, Mooney T, Cramm H. "Playing the system": structural factors potentiating mental health stigma, challenging awareness, and creating barriers to care for Canadian public safety personnel. Health (London). May 16, 2020;24(3):259-278. [ CrossRef ] [ Medline ]
- McCall HC, Beahm JD, Fournier AK, Burnett JL, Carleton RN, Hadjistavropoulos HD. Stakeholder perspectives on internet-delivered cognitive behavioural therapy for public safety personnel: a qualitative analysis. Can J Behav Sci. Jul 2021;53(3):232-242. [ CrossRef ]
- Hadjistavropoulos HD, McCall HC, Thiessen DL, Huang Z, Carleton RN, Dear BF, et al. Initial outcomes of transdiagnostic internet-delivered cognitive behavioral therapy tailored to public safety personnel: longitudinal observational study. J Med Internet Res. May 05, 2021;23(5):e27610. [ FREE Full text ] [ CrossRef ] [ Medline ]
- McCall H, Dear BF, Landry C, Beahm JD, Gregory J, Titov N, et al. Internet-delivered cognitive behavioural therapy for symptoms of PTSD among public safety personnel: initial outcomes of an open cohort preference trial of transdiagnostic and disorder-specific therapy. Internet Interv. Sep 2023;33:100656. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Hadjistavropoulos HD, McCall HC, Dear BF, Beahm JD, Carleton RN, Titov N. Outcomes of transdiagnostic internet-delivered cognitive behavioural therapy tailored to public safety personnel: a longitudinal observational study. J Anxiety Disord. Jun 2024;104:102861. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Kang M, Ragan BG, Park JH. Issues in outcomes research: an overview of randomization techniques for clinical trials. J Athl Train. 2008;43(2):215-221. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Sheehan DV. The Sheehan disability scales: the anxiety disease and how to overcome it. Mapi Research Trust. URL: https://eprovide.mapi-trust.org/instruments/sheehan-disability-scale [accessed 2024-04-29]
- Sharpe D, Cribbie RA. Analysis of treatment-control pre-post-follow-up design data. Quant Method Psychol. Feb 01, 2023;19(1):25-46. [ CrossRef ]
- Butcher NJ, Monsour A, Mew EJ, Chan AW, Moher D, Mayo-Wilson E, et al. Guidelines for reporting outcomes in trial reports: the CONSORT-outcomes 2022 extension. JAMA. Dec 13, 2022;328(22):2252-2264. [ CrossRef ] [ Medline ]
- Sunderland A, Findlay LC. Perceived need for mental health care in Canada: results from the 2012 Canadian Community Health Survey-Mental Health. Health Rep. Sep 2013;24(9):3-9. [ FREE Full text ] [ Medline ]
- Young C, Helis E, Williams D. Internet-delivered cognitive behavioural therapy for major depressive disorder and anxiety disorders: an environmental scan. Canadian Agency for Drugs and Technologies in Health. 2018. URL: https://www.cadth.ca/sites/default/files/pdf/ES0331_iCBT_for_Major_Depression_and_Anxiety.pdf [accessed 2024-04-29]
- Andersson G, Titov N, Dear BF, Rozental A, Carlbring P. Internet-delivered psychological treatments: from innovation to implementation. World Psychiatry. Feb 2019;18(1):20-28. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Donohue MC, Edland SD, Hu N. longpower: sample size calculations for longitudinal data. Cran R. URL: https://cran.r-project.org/web/packages/longpower/longpower.pdf [accessed 2024-04-29]
- Dear BF, Staples LG, Terides MD, Fogliati VJ, Sheehan J, Johnston L, et al. Transdiagnostic versus disorder-specific and clinician-guided versus self-guided internet-delivered treatment for social anxiety disorder and comorbid disorders: a randomized controlled trial. J Anxiety Disord. Aug 2016;42:30-44. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Berger T, Hämmerli K, Gubser N, Andersson G, Caspar F. Internet-based treatment of depression: a randomized controlled trial comparing guided with unguided self-help. Cogn Behav Ther. 2011;40(4):251-266. [ CrossRef ] [ Medline ]
- Andrews G, Basu A, Cuijpers P, Craske MG, McEvoy P, English CL, et al. Computer therapy for the anxiety and depression disorders is effective, acceptable and practical health care: an updated meta-analysis. J Anxiety Disord. Apr 2018;55:70-78. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Kroenke K, Spitzer RL, Williams JB. The PHQ-9: validity of a brief depression severity measure. J Gen Intern Med. Sep 2001;16(9):606-613. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Kroenke K, Spitzer RL, Williams JB, Löwe B. The patient health questionnaire somatic, anxiety, and depressive symptom scales: a systematic review. Gen Hosp Psychiatry. 2010;32(4):345-359. [ CrossRef ] [ Medline ]
- Manea L, Gilbody S, McMillan D. Optimal cut-off score for diagnosing depression with the Patient Health Questionnaire (PHQ-9): a meta-analysis. CMAJ. Feb 21, 2012;184(3):E191-E196. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Spitzer RL, Kroenke K, Williams JB, Löwe B. A brief measure for assessing generalized anxiety disorder: the GAD-7. Arch Intern Med. May 22, 2006;166(10):1092-1097. [ CrossRef ] [ Medline ]
- Blevins CA, Weathers FW, Davis MT, Witte TK, Domino JL. The posttraumatic stress disorder checklist for DSM-5 (PCL-5): development and initial psychometric evaluation. J Trauma Stress. Dec 2015;28(6):489-498. [ CrossRef ] [ Medline ]
- Bovin MJ, Marx BP, Weathers FW, Gallagher MW, Rodriguez P, Schnurr PP, et al. Psychometric properties of the PTSD checklist for diagnostic and statistical manual of mental disorders-fifth edition (PCL-5) in veterans. Psychol Assess. Nov 2016;28(11):1379-1391. [ CrossRef ] [ Medline ]
- Wortmann JH, Jordan AH, Weathers FW, Resick PA, Dondanville KA, Hall-Clark B, et al. Psychometric analysis of the PTSD Checklist-5 (PCL-5) among treatment-seeking military service members. Psychol Assess. Nov 2016;28(11):1392-1403. [ CrossRef ] [ Medline ]
- Leontjevas R, de Beek WO, Lataster J, Jacobs N. Resilience to affective disorders: a comparative validation of two resilience scales. J Affect Disord. Oct 2014;168:262-268. [ CrossRef ] [ Medline ]
- Smith BW, Dalen J, Wiggins K, Tooley E, Christopher P, Bernard J. The brief resilience scale: assessing the ability to bounce back. Int J Behav Med. 2008;15(3):194-200. [ CrossRef ] [ Medline ]
- Diener E, Wirtz D, Biswas-Diener R, Tov W, Kim-Prieto C, Choi DW, et al. New measures of well-being. In: Diener E, editor. Assessing Well-Being: The Collected Works of Ed Diener. Cham, Switzerland. Springer; 2009:247-266.
- Diener E, Wirtz D, Tov W, Kim-Prieto C, Choi D, Oishi S, et al. New well-being measures: short scales to assess flourishing and positive and negative feelings. Soc Indic Res. May 28, 2009;97(2):143-156. [ CrossRef ]
- Miller SD, Duncan BL, Johnson LD, Reynolds LR, Brown J. The session rating scale: preliminary psychometric properties of a “working” alliance measure. J Brif Ther. 2000;3(1):3-12. [ FREE Full text ]
- Campbell A, Hemsley S. Outcome rating scale and session rating scale in psychological practice: clinical utility of ultra-brief measures. Clin Psychol. Mar 11, 2009;13(1):1-9. [ CrossRef ]
- Duncan BL, Miller SD, Sparks JA, Claud DA, Reynolds LR, Brown J, et al. The session rating scale: preliminary psychometric properties of a “working” alliance measure. J Brif Ther. 2003;3(1):3-12. [ FREE Full text ]
- Cavanagh K, Herbeck Belnap B, Rothenberger SD, Abebe KZ, Rollman BL. My care manager, my computer therapy and me: the relationship triangle in computerized cognitive behavioural therapy. Internet Interv. Mar 2018;11:11-19. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Devilly GJ, Borkovec TD. Psychometric properties of the credibility/expectancy questionnaire. J Behav Ther Exp Psychiatry. Jun 2000;31(2):73-86. [ CrossRef ] [ Medline ]
- Saunders JB, Aasland OG, Babor TF, de la Fuente JR, Grant M. Development of the alcohol use disorders identification test (AUDIT): WHO collaborative project on early detection of persons with harmful alcohol consumption--II. Addiction. Jun 24, 1993;88(6):791-804. [ CrossRef ] [ Medline ]
- Berman AH, Bergman H, Palmstierna T, Schlyter F. Evaluation of the drug use disorders identification test (DUDIT) in criminal justice and detoxification settings and in a Swedish population sample. Eur Addict Res. 2005;11(1):22-31. [ CrossRef ] [ Medline ]
- McCall HC, Sison AP, Burnett JL, Beahm JD, Hadjistavropoulos HD. Exploring perceptions of internet-delivered cognitive behaviour therapy among public safety personnel: informing dissemination efforts. Int J Environ Res Public Health. Aug 19, 2020;17(17):6026. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Titov N, Dear BF, Staples LG, Bennett-Levy J, Klein B, Rapee RM, et al. MindSpot clinic: an accessible, efficient, and effective online treatment service for anxiety and depression. Psychiatr Serv. Oct 2015;66(10):1043-1050. [ CrossRef ] [ Medline ]
- Hadjistavropoulos HD, Peynenburg V, Thiessen DL, Nugent M, Karin E, Staples L, et al. Utilization, patient characteristics, and longitudinal improvements among patients from a provincially funded transdiagnostic internet-delivered cognitive behavioural therapy program: observational study of trends over 6 years. Can J Psychiatry. Mar 12, 2022;67(3):192-206. [ FREE Full text ] [ CrossRef ] [ Medline ]
- McCall HC, Hadjistavropoulos HD, Loutzenhiser L. Reconsidering the ethics of exclusion criteria in research on digital mental health interventions. Ethics Behav. Oct 31, 2019;31(3):171-180. [ CrossRef ]
- Harvey LA. Statistical testing for baseline differences between randomised groups is not meaningful. Spinal Cord. Oct 4, 2018;56(10):919. [ CrossRef ] [ Medline ]
- Gupta SK. Intention-to-treat concept: a review. Perspect Clin Res. Jul 2011;2(3):109-112. [ FREE Full text ] [ CrossRef ] [ Medline ]
- McNeish D. Small sample methods for multilevel modeling: a colloquial elucidation of REML and the Kenward-Roger correction. Multivariate Behav Res. Jul 17, 2017;52(5):661-670. [ CrossRef ] [ Medline ]
- Field AP. Discovering Statistics Using IBM SPSS Statistics. 5th edition. Thousand Oaks, CA. Sage Publications; 2018.
- Hair JF, Black WC, Babin BJ, Anderson RE. Multivariate Data Analysis. 7th edition. Harlow, UK. Pearson; 2014.
- Edmonds M, Hadjistavropoulos HD, Schneider LH, Dear BF, Titov N. Who benefits most from therapist-assisted internet-delivered cognitive behaviour therapy in clinical practice? predictors of symptom change and dropout. J Anxiety Disord. Mar 2018;54:24-32. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Beatty L, Binnion C. A systematic review of predictors of, and reasons for, adherence to online psychological interventions. Int J Behav Med. Dec 2016;23(6):776-794. [ CrossRef ] [ Medline ]
- Constantino MJ, Arnkoff DB, Glass CR, Ametrano RM, Smith JZ. Expectations. J Clin Psychol. Feb 2011;67(2):184-192. [ CrossRef ] [ Medline ]
- Thompson-Hollands J, Bentley KH, Gallagher MW, Boswell JF, Barlow DH. Credibility and outcome expectancy in the unified protocol: relationship to outcomes. J Exp Psychopathol. Mar 30, 2014;5(1):72-82. [ CrossRef ]
- Haller K, Becker P, Niemeyer H, Boettcher J. Who benefits from guided internet-based interventions? a systematic review of predictors and moderators of treatment outcome. Internet Interv. Sep 2023;33:100635. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Lambert MJ. Psychotherapy outcome research: implications for integrative and eclectical therapists. In: Norcross JC, Goldfried MR, editors. Handbook of Psychotherapy Integration. New York, NY. Basic Books; 1992:94-129.
- Elo S, Kyngäs H. The qualitative content analysis process. J Adv Nurs. Apr 2008;62(1):107-115. [ CrossRef ] [ Medline ]
- Berger R. Now I see it, now I don’t: researcher’s position and reflexivity in qualitative research. Qual Res. Jan 03, 2013;15(2):219-234. [ CrossRef ]
- The improving access to psychological therapies manual: appendices and helpful resources. National Collaborating Centre for Mental Health. 2019. URL: https://www.england.nhs.uk/wp-content/uploads/2018/06/iapt-manual-appendices-and-helpful-resources-v3.pdf [accessed 2020-12-15]
- Titov N, Dear BF, Staples LG, Bennett-Levy J, Klein B, Rapee RM, et al. The first 30 months of the mindspot clinic: evaluation of a national e-mental health service against project objectives. Aust N Z J Psychiatry. Dec 2017;51(12):1227-1239. [ CrossRef ] [ Medline ]
- Titov N, Dear BF, Nielssen O, Wootton B, Kayrouz R, Karin E, et al. User characteristics and outcomes from a national digital mental health service: an observational study of registrants of the Australian MindSpot Clinic. Lancet Digit Health. Nov 2020;2(11):e582-e593. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Byllesby BM, Dickstein BD, Chard KM. The probability of change versus dropout in veterans receiving cognitive processing therapy for posttraumatic stress disorder. Behav Res Ther. Dec 2019;123:103483. [ CrossRef ] [ Medline ]
- Richards D, Richardson T. Computer-based psychological treatments for depression: a systematic review and meta-analysis. Clin Psychol Rev. Jun 2012;32(4):329-342. [ CrossRef ] [ Medline ]
- Beahm JD, McCall HC, Carleton RN, Titov N, Dear B, Hadjistavropoulos HD. Insights into internet-delivered cognitive behavioural therapy for public safety personnel: exploration of client experiences during and after treatment. Internet Interv. Dec 2021;26:100481. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Ljótsson B, Falk L, Vesterlund AW, Hedman E, Lindfors P, Rück C, et al. Internet-delivered exposure and mindfulness based therapy for irritable bowel syndrome--a randomized controlled trial. Behav Res Ther. Jun 2010;48(6):531-539. [ CrossRef ] [ Medline ]
- O'Mahen HA, Woodford J, McGinley J, Warren FC, Richards DA, Lynch TR, et al. Internet-based behavioral activation--treatment for postnatal depression (Netmums): a randomized controlled trial. J Affect Disord. Sep 25, 2013;150(3):814-822. [ CrossRef ] [ Medline ]
- Goldstein NJ, Cialdini RB, Griskevicius V. A room with a viewpoint: using social norms to motivate environmental conservation in hotels. J Consum Res. Oct 01, 2008;35(3):472-482. [ CrossRef ]
- Carlbring P, Lindqvist K, Mechler J, Philips B. Transdiagnostic self-help treatments for anxiety and/or depression: a full-factorial RCT investigating psychotherapeutic modalities, discussion boards and treatment length. In: Proceedings of the 11th Scientific Meeting on International Society for Research on Internet Interventions. 2022. Presented at: ISRII '22; September 18-21, 2022:1-19; Pittsburgh, PA. URL: https://www.diva-portal.org/smash/record.jsf?pid=diva2%3A1700061&dswid=-46
Abbreviations
| Brief Resilience Scale |
| Credibility/Expectancy Questionnaire |
| Consolidated Standards of Reporting Trials |
| digital mental health intervention |
| Flourishing Scale |
| Generalized Anxiety Disorder–7 |
| internet-delivered cognitive behavioral therapy |
| multilevel modeling |
| PTSD Checklist for DSM-5 |
| Patient Health Questionnaire–9 |
| public safety personnel |
| posttraumatic stress disorder |
| Session Rating Scale |
Edited by A Mavragani; submitted 19.04.24; peer-reviewed by L Yang; comments to author 10.06.24; revised version received 26.06.24; accepted 22.07.24; published 14.08.24.
©Hugh C McCall, Heather D Hadjistavropoulos. Originally published in the Journal of Medical Internet Research (https://www.jmir.org), 14.08.2024.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work, first published in the Journal of Medical Internet Research (ISSN 1438-8871), is properly cited. The complete bibliographic information, a link to the original publication on https://www.jmir.org/, as well as this copyright and license information must be included.
IMAGES
COMMENTS
Behavioural genetics articles from across Nature Portfolio. Behavioural genetics is the interdisciplinary effort to establish causal links between genes and animal (including human) behavioural ...
Celebrating a Century of Research in Behavioral Genetics. A century after the first twin and adoption studies of behavior in the 1920s, this review looks back on the journey and celebrates milestones in behavioral genetic research. After a whistle-stop tour of early quantitative genetic research and the parallel journey of molecular genetics ...
In the context of current concerns about replication in psychological science, we describe 10 findings from behavioral genetic research that have robustly replicated. These are 'big' findings, both in terms of effect size and potential ...
For fields such as social and behavioral genomics that are shaped by an ugly history and uncertain future, socially and ethically responsible research and research communication are crucial. FoGS ...
Overview. Behavior Genetics is a leading journal concerned with the genetic analysis of behavioral traits. The journal offers the most current original research on the inheritance and evolution of behavioral characteristics in humans and other species. Contributions focus on both the application of various genetic perspectives to the study of ...
Read the latest Research articles in Behavioural genetics from Nature
Behavioral genetics and behavioral genomics as complementary approaches. Both behavioral genetics and behavioral genomics investigate the link between genes and behavioral variation. Behavioral genetics, as exemplified by Peromyscus burrowing, seeks to connect inherited genetic variation to behavioral variation.
However, polygenic scores are useful for assessing the genetic contribution of a trait to variation in (relatively) small groups of individuals, the sort of size collected in social science research rather than the hundreds of thousands that geneticists amass for genome-wide association studies.
Psychiatric genetics has made substantial progress in the last decade, providing new insights into the genetic etiology of psychiatric disorders, and paving the way for precision psychiatry, in which individual genetic profiles may be used to personalize ...
Behavior Genetics is a leading journal concerned with the genetic analysis of behavioral traits. The journal offers the most current original research on ...
Behavior genetics is the study of the manner in which genetic variation affects psychological phenotypes (traits), including cognitive abilities, personality, mental illness, and social attitudes. In a seminal article published in this journal, Turkheimer (2000) noted three robust empirical regularities that had by then emerged from the literature on behavior genetics. He dubbed these ...
In this review I will focus on a few key issues facing contemporary behavior genetics. Behavior genetics is, in principle, not very different from other subfields of genetics: It is strongly multidisciplinary and interdisciplinary, with contributions from ethology, psychology, neuroscience, ecology, psychiatry, etc., and focuses on a specific ...
Human behavioral genetic research aimed at characterizing the existence and nature of genetic and environmental influences on individual differences in cognitive ability, personality and interests, and psychopathology is reviewed. Twin and adoption studies indicate that most behavioral characteristics are heritable.
Blaming Our Genes: The Heritability of Behavior. When it comes to behavior, genetics may play a larger role than you think. It's easy to accept that human disorders such as phenylketonuria or cystic fibrosis or Huntington's disease have a wholly genetic basis. And you likely have no problem believing that your risk of being afflicted with ...
Behavioural genetics examines the underlying factors associated with individual differences in behaviours and capacities. In this Primer, Willoughby et al. discuss the methods used in behavioural ...
This article provides an overview of the methods, assumptions, and key findings of behavioral genetics methodology for family researchers with a limited background. We discuss how family researchers can utilize and contribute to the behavioral genetics field, particularly in terms of conducting rese …
As behavioral genetics enters a second century, the field's oldest research method remains both relevant and controversial.
A century after the first twin and adoption studies of behavior in the 1920s, this review looks back on the journey and celebrates milestones in behavioral genetic research. After a whistle-stop tour of early quantitative genetic research and the parallel journey of molecular genetics, the travelogue focuses on the last fifty years. Just as quantitative genetic discoveries were beginning to ...
Recent victimology research, however, has shown that genetic and evolutionary forces are connected to the risk of victimization. The current study expands on these findings by examining whether genetic influences differentially explain victimization in males and females.
Rare and common variants contribute incrementally to a continuum of behavioral and developmental traits, of which the severe tail results in ASD diagnosis (76, 77), and the total genetic load sufficient for diagnosis, or threshold, can be reached through differing combinations of rare and common variation (31, 78-80).
This Article discusses a behavioral genetic perspective that provides insight into the biological factors that influence family relationships. Part II presents a brief overview of the research methods used to understand both genetic and environmental influences on human behavior.
Decision Making ; News ; Scientists Capture Clearest Glimpse of How Brain Cells Embody Thought Recordings from thousands of neurons reveal how a person's brain abstractly represents acts of reasoning.
The connection between chronic psychological stress and the onset of various diseases, including diabetes, HIV, cancer, and cardiovascular conditions, is well documented. This review synthesizes current research on the neurological, immune, hormonal, and genetic pathways through which stress influences disease progression, affecting multiple body systems: nervous, immune, cardiovascular ...
Research is shedding light on different chromosomal make-ups and what advantages they may bring to sport.
It's hoped the findings lead to more humane horse training regimes and improvements to welfare.
Harden and Koellinger discuss the goals, methods and challenges of social science genetics, which aims to unravel the genetic underpinnings of individual differences in social, behavioural and ...
Researchers have found that molecules and microorganisms both inside and outside our bodies are going through dramatic changes, first around age 44 and again at 60.
This article provides an overview of the methods, assumptions, and key findings of behavioral genetics methodology for family researchers with a limited background. We discuss how family researchers can utilize and contribute to the behavioral genetics field, particularly in terms of conducting research that seeks to explain shared ...
A new study has identified new genetic variants associated with normal pressure hydrocephalus (NPH). The results provide new insights into the genetic background of NPH and provide a basis for ...
Background: Internet-delivered cognitive behavioral therapy (ICBT) is an effective and accessible treatment for various mental health concerns. ICBT has shown promising treatment outcomes among public safety personnel (PSP), who experience high rates of mental health problems and face barriers to accessing other mental health services. Client engagement and clinical outcomes are better in ICBT ...